
Úvod

Tyto materiály byly inovovány za podpory OPVK proejktu EnviMod (http://envimod.fzp.ujep.cz)
CZ.1.07/2.2.00/28.0205 „Modernizace výuky technických a přírodovědných oborů na UJEP se zaměřením na problematiku ochrany životního prostředí“ – ENVIMOD
Tyto materiály byly inovovány za podpory OPVK proejktu EnviMod (http://envimod.fzp.ujep.cz)
Učební text.
Mikrobiologie je věda, zabývající se zejména mikroskopickými organismy, mezi které patří např. bakterie, viry nebo mikroskopické houby. Název mikrobiologie pochází z řečtiny, mikros znamená malý, bios znamená život a logos znamená slovo. Historicky se mikrobiologie zabývala zejména těmi organismy, které nebyly vidět pouhým okem, moderní mikrobiologie už se tak striktního vymezení nedrží.
Mikrobiologie má mnoho podoborů, např. virologie zkoumá viry (nebuněčné parazity buněk), bakteriologie bakterie, algologie prokaryotické řasy, mykologie zejména houby atd. Mikrobiologie jako biologická věda využívá poznatků dalších věd, např. molekulární biologie, ekologie či biochemie. Naopak mnoho aplikovaných oborů využívá mikrobiologických poznatků, např. biotechnologie, lékařská mikrobiologie nebo mikrobiologie potravin.
Historii mikrobiologie můžeme datovat už do starověku. Např. římský filozof Lucretius se domníval, že nemoci jsou způsobovány neviditelnými organismy. Těžko říci, jak se dobral tohoto závěru, nicméně je obecně správný. Středověk pak znamenal pro mikrobiologii, stejně jako pro většinu ostatních věd období temna. Z hlediska mikrobiologického je tak středověk zajímavý jen častými výskyty epidemií nejrůznějších smrtelných chorob, nejničivější byly zejména mor a cholera.
Za začátek novodobé mikrobiologie lze považovat práci Roberka Hooka „Micrographia“ (1665) ve které poprvé použil termínu buňka v dnes chápaném významu základní jednotky živé hmoty. Skutečnou revoluci v mikrobiologii pak přinesl vynález mikroskopu Antonym van Leeuwenhoekem v roce 1673. Leeuwenhoek byl pravděpodobně prvním člověkem, který spatřil na vlastní oči bakterie. Od tohoto okamžiku se začala mikrobiologie rozvíjet jako skutečná věda, která shromažďovala, utřiďovala a zobecňovala získané poznatky. Z doby po Leuwenhoekovi pak pocházejí nejznámější mikrobiologická jména minulosti. Ferdinand Cohn (1828-1898) navrhl první systém klasifikace bakterií a objevil bakteriální spory. Jeho současník Louis Pasteur (1822-1895) se zapsal do dějin mikrobiologie např. objevem techniky likvidace bakterií teplem, která dodnes nese jeho jméno – pasterace. Pomocí pasterace se mu také podařilo vyvrátit do té doby uznávanou teorii samoplození a upevnit tak jeden základní biologický zákon, že živé vzniká vždy jen z živého. Pasteurův přínos mikrobiologii byl i v oblasti metodické, zavedl např. očkování mikroorganismů do dnes používaným způsobem.
Další mohutnou vlnou rozvoje mikrobiologie byl přelom 19. a 20. století. V roce 1892 popsal Dmitrij Ivanovský první virus (tabákové mozaiky), čímž fakticky založil další podobor mikrobiologie. Do tohoto období spadá pravděpodobně i nejslavnější mikrobiologický objev všech dob. V roce 1929 objevil Alexander Fleming první antibiotikum – penicilin. Milióny lidí vděčí penicilinu za svůj život, farmaceutické firmy na něm vydělaly miliardy dolarů, paradoxem ale zůstává, že za objevem stojí zejména Flemingova nepořádnost. Kdyby Flemingovi v průběhu dovolené nezplesnivěly misky se stafylokoky, možná bychom na tento objev ještě dlouho čekali.
Začátek 20. století také přinesl významné objevy na poli biochemie. Biochemici se pomalu přestával zabývat jen složením živé hmoty, ale začali se intenzivně zabývat také pochody, které v živých organismech probíhají. Intenzivně byly zkoumány zejména enzymy, byly objeveny první biochemické dráhy a také zkoumána vnitřní struktura buněk.
Skutečnou revoluci, která biologické vědy opanuje dodnes, znamenal objev struktury deoxyribonukleové kyseliny (DNA) Jamesem Watsonem, Francisem Crickem a Mauricem Wilkinsonem (1953). Dvojšroubovicový model otevřel vědcům oči a vysvětlení základních genetických pochodů na úrovni molekul na sebe nenechalo dlouho čekat. V roce 1966 byl pak kompletně rozluštěn genetický kód. Rozvoj genetiky vedl i ke snaze využít poznatků k zásahům do genetického vybavení. V roce 1973 tak spatřila světlo světa první geneticky modifikovaná bakterie Escherichia coli, nesoucí v sobě žabí geny. V roce 1995 byl úspěšně dokončen první projekt sekvenace kompletního genomu bakterie Haemophilus influenzae. Dnes je znám kompletní genom u stovek bakterií.
Mikroorganismy patří mezi tzv. živé soustavy. Ty se od neživých soustav odlišují několika základními charakteristikami:
Někteří autoři uvádějí i další charakteristiky živé hmoty, např. obsah bílkovin a nukleových kyselin (které nacházíme u všech dosud známých živých organismů) nebo buněčná stavba těla. Akademické spory se vedou o zařazení virů, které nejsou schopné samostatného rozmnožování a napadají za tímto účelem hostitelské buňky.
Mikroorganismy, ačkoliv mikroskopické, zaujímají významné až dominantní postavení v životním prostředí a mají v něm nezastupitelné úlohy.
Mikroorganismy jsou schopnými rozkladači zejména organické hmoty rozkládající např. těla jiných organismů (mrtvá i živá), jejich odumřelé součásti (např. listy ze stromu), látky vyloučené jinými organismy (trus apod.), mnohé horniny a nerosty nebo látky vytvořené člověkem.
Mikroorganismy jsou důležitými producenty chemických látek. Kromě vlastní biomasy produkují např. látky určené k boji proti jiným organismům (antibiotika, toxiny apod.), ochranné látky (např. polysacharidová pouzdra), enzymy katalyzující rozklad látek či odpadní produkty metabolismu, často dále využitelné jinými organismy či člověkem (ethanol, vodík, methan apod.).
Svou činností mají mikroorganismy nezastupitelnou úlohu při tvorbě půdy, čištění vod a v tzv. geochemických cyklech, tj. koloběhu základních chemických prvků v přírodě.
Mikroorganismy jsou součástí přirozených společenstev organismů v přírodě. Jejich vztahy k ostatním členům společenstva mohou být pozitivní, neutrální i negativní. Mnohé mikroorganismy žijí v symbióze s makroorganismy a toto spojení bývá často výhodné či dokonce nezbytně nutné.
V podobném vztahu jsou mikroorganismy i vůči člověku. Člověk hostí ve svém těle bilióny a trilióny bakterií, bez kterých by např. nebyl schopen správně trávit potravu. Mikroorganismy ale ovlivňují člověka i v jeho činnosti. Člověk mikroorganismy využívá ke svému prospěchu jako producenty žádaných látek, při likvidacích odpadu nebo produkci potravin. Naopak mikroorganismy člověka v mnoha ohledech škodí, ať už jako patogeny způsobující choroby či rozkladači potravin a jiných užitečných výrobků.
Výskyt organismů se dá charakterizovat velice jednoduše: mikroorganismy jsou prakticky všude. Tzv. aseptické prostředí (tj. prostředí bez mikroorganismů) se na Zemi přirozeně vyskytuje velmi zřídka a obvykle je jen dílem člověka. Příkladem jsou aseptická prostředí v nemocnicích a vědeckých ústavech a pak lokality s opravdu extrémními podmínkami (rozžhavená láva apod.).
Variabilita a odolnost mikroorganismů je podstatně větší než makroorganismů a mnohé mikroorganismy osidlují i prostředí, kde panují z lidského pohledu extrémní podmínky, např. polární kraje, jícny podmořských sopek či extrémně slané vody, např. Mrtvé moře. Všude tam nacházíme unikátní mikroorganismy dobře přizpůsobené pro dané prostředí.
I vzhledem ke své velikosti se mikroorganismy počítají obvykle v tisících, miliónech či miliardách, viz. tabulka:
|
Prostředí |
Orientační množství bakterií |
|
Stolice zdravého člověka |
109 /g |
|
Zemědělská půda |
108 /g |
|
Bachorová šťáva přežvýkavců |
1010 / ml |
|
Lidská slina |
108 /ml |
|
Jezera a rybníky |
106 /ml |
|
Jogurt |
>109 /g |
|
Pitná voda |
102 /ml |
|
Vzduch |
103 /m3 |
Dnes nejrozšířenější názor na vznik života je ten, že život na Zemi vznikl samovolně z neživé hmoty a postupným vývojem se rozvinul až do dnešní bohaté podoby. Tato evoluční teorie má nicméně mnoho nejasností, bílých míst a fakty málo podložených hypotéz. I díky nemalému rozšíření nejrůznějších náboženství tak existuje mnoho odpůrců evoluční teorie. Kreacionisté věří, že život byl stvořen bohem, eternitisté věří tomu, že život je věčný a vzniknout tak ani nemusel. Neobvyklý není ani ten názor, že život vznikl na jiné planetě a byl na Zemi zanesen. Většina vědců nicméně věří, že nejasnosti se podaří časem vysvětlit a tato víra byla v minulosti mnohokrát podpořena novými objevy. Nové objevy pak také vedou k upřesnění časového vývoje, následující řádky proto možná nebudou za pár let platit.
Země vznikla pravděpodobně přibližně před 4,5 miliardami let a byla výrazně jiná než dnes. Atmosféra byla pravděpodobně redukční a sestávala zejména z methanu, amoniaku, vodíku, vodních par, dusíku, amoniaku a oxidu uhličitého. Vlivem slunečního záření (zejména ultrafialového) a vlivem častých bouří docházelo v atmosféře k chemickým reakcím, které daly vzniknout složitějším chemickým látkám, např. aminokyselinám (součást bílkovin), purinům (součást DNA) a mnohým dalším. Tuto fázi vývoje života nazýváme obvykle chemickou evolucí a její stěžejní reakce se podařilo nasimulovat v laboratořích. Kumulací a vysycháním těchto jednoduchých organických látek vznikaly pravděpodobně polymery (jakési prabílkoviny a pranukleové kyseliny), které se za určitých nejasných okolností shlukovaly za vzniku přibližně kulatých útvarů. S velmi malou ale nenulovou pravděpodobností byl některý z těchto útvarů natolik organizovaný, že měl schopnost rozmnožování a dal tak fakticky vzniknout první živé buňce. Primitivní život byl pravděpodobně založen zejména na ribonukleové kyselině (RNA), která vykonávala nejen funkci informační molekuly, ale též funkci stavební a katalytickou. Pravděpodobným pozůstatkem této fáze praživota jsou funkční ribozomální a transferové RNA, které hrají hlavní roli v syntéze bílkovin.
Biologická evoluce začíná vznikem genetického kódu a proteosyntézy. Genetický kód všech dnešních organismů je univerzální (tj. pro všechny stejný) a je tedy zjevné, že mají jednoho společného předka. Vývoj od tohoto předka se pak ubíral dvěma hlavními směry, první větev zůstala na nižší buněčné úrovni a dala vzniknout dnešním bakteriím. Druhá větev se později rozdělila na další dvě. Z první se vyvinuly organismy jejichž stavba buněk zůstala primitivní podobná bakteriím, ale s některými pokročilejšími vlastnostmi. Tyto organismy byly nazvány archea. Druhá větev se vyvinula k pokročilejší stavbě buňky a dala vzniknout dnešním eukaryotickým organismům – rostlinám, živočichům, houbám a prvokům.
Společný vývoj archeí a eukaryot je doložen např.
genetickými mechanismy, které jsou u obou skupin velice podobné a proti genetickým
mechanismům bakterií výrazně pokročilejší. Samostatný vývoj archeí je pak
doložen některými unikátními metabolickými drahami (např. methanogenezí) či
unikátními prvky ve stavbě buněk (např. buněčné membrány).
Otázkou zůstává, jakým způsobem se vyvinula pokročilá eukaryotická buňka. Tak je totiž, na rozdíl od bakteriálních a archeálních buněk, vnitřně rozdělená membránami na různé kompartmenty a organely. Některé z těchto organel (mitochondrie a chloroplasty) jsou přitom částečně autonomní a mají i vlastní genetickou informaci (podobnou bakteriální). Dnes se předpokládá tzv. endosymbiotická teorie, podle které pohltila primitivní eukaryotická buňka purpurové bakterie. Tato symbióza posléze přešla v závislost a specializaci a z původní purpurové bakterie se stala mitochondrie. K tomuto jevu došlo pravděpodobně několikrát. Předpokládá se, že rostlinné chloroplasty vznikly podobným mechanismem pohlcením zelené fotosyntetizující bakterie. Dnešní rostlinná buňka (nesoucí jak mitochondrie, tak chloroplasty) pravděpodobně vznikla splynutím dvou buněk, jedné nesoucí chloroplasty, druhé nesoucí mitochondrie.
Základem každé vědy je systematika získaných poznatků a biologické vědy nejsou pochopitelně žádnou výjimkou. Biologové se odjakživa snažili známé organismy nějakým způsobem zatřídit do systému. Vznikl tak podobor biologie nazývaný taxonomie. Základy dnes používaného taxonomického systému položil už v 18. století švédský biolog Carl Linné. Ten ve svém díle Systematica nature (Soustava přírody) z roku 1735 stručně popsal všechny tehdy známé biologické druhy a každému přiřadil dvojslovný název – rodové a druhové jméno. Vznikla tak tzv. binomická nomenklatura, kterou biologie využívá dodnes. Rodové a druhové jméno se v odborných publikacích píše tradičně proloženě (kurzívou).
Základní taxonomickou jednotkou je druh. U vyšších organismů je jeho vymezení jednoduché, jedinci stejného druhu jsou ty, kteří jsou schopni se vzájemně rozmnožovat za vzniku plodního potomstva. V mikrobiologii ale tato definice naráží, protože naprostá většina mikroorganismů se rozmnožuje pouze nepohlavně, popř. střídavě pohlavně a nepohlavně. U těchto organismů (zejména bakterií) proto dáváme přednost definici druhu založené na dostatečné vzájemné podobnosti organismů, obvykle se uvažuje 95% shoda. U bakterií pak narážíme na další problém a tím je poměrně velká morfologická uniformita. Tvary bakteriálních buněk nejsou příliš rozmanité, jejich pozorování je vzhledem k velikosti i s obyčejným mikroskopem obtížné a pro taxonomické hodnocení se příliš nehodí. Proto se v mikrobiologii používají zejména chemické a metabolické charakteristiky. V posledních letech se pak dostávají do popředí zejména charakteristiky založené na genetické informaci a sekvencích DNA a RNA. Ty mají mnoho výhod, např. že odrážejí vývojovou (fylogenetickou) příbuznost nebo že nezávisí na fyziologickém stavu organismů.
Taxonomie se ale nespokojuje jen s určením druhu, ale snaží se zatřídit organismy do větších hierarchických skupin, tzv. taxonů. Každý taxon je založen na dostatečné míře vzájemné podobnosti jeho členů a zároveň na dostatečné míře odlišnosti těchto členů od členů patřících do jiného taxonu. Míru podobnosti organismů můžeme matematicky vyjádřit formou čtyřpolní tabulky:
|
|
počet vlastností, které druh |
1. druh |
|
|
2. druh |
má |
nemá |
|
|
má |
a |
b |
|
|
nemá |
c |
d |
|
Políčka označená jako a, b, c, d se vyplní počty znaků, které se vyskytují u obou organismů (a), které pouze u jednoho (b resp. c) a které se nevyskytují u žádného organismu. Ze čtyřpolní tabulky je pak možné vypočítat koeficient podobnosti, např. tzv. koeficient prosté shody (simple matching coefficient):
SSM = (a+d) / (a+b+c+d)
popř. tzv. Jaccardův koeficient, který nezahrnuje znaky nevyskytující se ani u jednoho porovnávaného druhu:
SSM = a / (a+b+c)
Oba koeficienty dosahují hodnot <0-1>, čím vyšší hodnota, tím vyšší vzájemná podobnost.
Na základě koeficientů podobnosti je pak možné sestavovat tzv. stromové grafy, ve kterých leží podobné organismy blíže u sebe a jejichž větvení odpovídá vyšším taxonům (tzv. fenonům).
Při porovnávání podobnosti jednotlivých organismů je možné využít jakýchkoliv pozorovatelných vlastností organismů, čím jich je více, tím větší je přesnost takového porovnání. Tradičně se k tomuto účelu využívají zejména morfologické (tvarové) znaky (tvar těla, barva apod.). Rozvoj geneticky dal nicméně biologům do ruky novou a podstatně mocnější zbraň – porovnávání genetických sekvencí. Zatímco u morfologických znaků už bývá velmi problematické a hlavně pracné správné určení už několik desítek znaků, genetické sekvence nabízejí o několik řádů bohatší informaci. Navíc se předpokládá, že náhodné změny genetické informace (mutace) jsou hlavní hybnou silou evoluce nových znaků a vlastností. Fenotypové (tj. vnější) projevy těchto mutací jsou pak podrobeny tlaku prostředí (selekce), výhodné jsou zachovány (fixovány), nevýhodné naopak obvykle eliminovány. Příbuznější organismy proto budou mít vzájemně podobnější genetické sekvence.
Jako nejvhodnější část genetické informace pro určení evoluční příbuznosti se ukázala ribozomální RNA. Ribozómy nacházíme u všech organismů od bakterií až po člověka a z těchto sekvencí je tedy možné sestrojit skutečně univerzální fylogenetický strom. Navíc mají ribozomální RNA v ribozómech stavební funkci, mutace v této oblasti jsou proto veskrze neutrální a selekce prakticky nenastává.
Univerzální fylogenetcický strom založený na 16S RNA (resp. 18SRNA u eukaryot) je uveden na Obr. 1. Z něj vychází základní evoluční linie nazvané nově domény (Bacteria, Archea a Eukarya) i jejich další větvení.
Z evoluční příbuznosti vychází také moderní taxonomický systém. Taxonomové se snaží vyšší taxony uspořádávat tak, aby byly tzv. monofyletické, tj. aby v každém taxonu byli pouze ty druhy, které mají jednoho společného předka. Příkladem takovýchto taxonů jsou např. tři základní eukaryotické říše (rostliny, živočichové a houby). Monofyletické taxony jsou považovány za přirozené. Opakem jsou umělé tzv. polyfyletické taxony, který sdružují organismy s více předky. Příkladem může být umělá říše Prvoci (Protozoa), která zahrnuje menší nezávislé eukaryotické vývojové linie. Je samozřejmě pravda, že všichni prvoci mají jednoho společného předka (praeukaryotický organismus), nicméně takovýto taxon by odpovídal všem eukaryotům a nezahrnoval už by jen prvoky.
Hlavní taxony používané v biologii uvádí tabulka:
|
Česky |
Latinsky |
Anglicky |
Přípona |
|
doména |
regio |
domain |
|
|
říše |
regnum |
kingdom |
|
|
kmen |
phyllum |
phylum |
|
|
oddělení |
division |
division |
|
|
třída |
classis |
class |
|
|
řád |
ordo |
order |
-ales |
|
čeleď |
familia |
family |
-aceae |
|
rod |
genus |
genus |
|
|
druh |
species |
species |
|
Objektivně je ale třeba dodat, že v mikrobiologii nenašly tyto taxony velkého uplatnění. Díky absenci pohlavního rozmnožování u většiny mikroorganismů bývá totiž obvykle problematické určitě správně rod a druh, natož pak vyšší taxony. Systematická mikrobiologie se proto obvykle drží spíše tradičního, ale praktičtější členění založeného zejména na biochemii a morfologii (viz kapitola 12).
V mikrobiologii mají naopak daleko větší uplatnění nižší taxony. Zejména u bakterií se často definuje tzv. kmen, který představuje bližší charakteristiku zástupců druhu. Tato charakteristika může být např. místo, ze kterého byl daný kmen izolován. Zástupci kmene pak obvykle představují potomky (klony) jediné konkrétní bakterie. Jiná charakteristika může být např. konkrétní mutace nebo přítomnost či absence některých genů. Kmen se obvykle vyznačuje kratší sekvencí písmen, kterou určuje ten, kdo kmen izoloval a který se nepíše kurzívou. Přítomnost či absence genů se pak obvykle vyznačuje krátkými třípísmenými zkratkami biochemických drah s označením + či -, které se naopak kurzívou píší. Ilustračním příkladem mohou být čtyři kmeny bakterie Pseudomonas fluorescens některé geneticky modifikované:
Pseudomonas fluorescens 5R nah+ sal+ lux-
Pseudomonas fluorescens 5RL nah+ sal- lux+
Pseudomonas fluorescens HK9 nah+ sal+ lux-
Pseudomonas fluorescens HK44 nah+ sal+ lux+
První kmen představuje přirozený izolát který má funkční geny pro odbourávání naftalenu (nah+) i salicylové kyseliny (sal+), ale nemá geny pro luminiscenci. Druhý kmen představuje genetickou modifikaci předchozího kmenu, do kterého byly přidány luminiscenční geny (lux+). Vložení těchto genů ale poškodilo geny pro odbourávání salicylové kyseliny a proto je tento kmen sal-. Třetí kmen je pak opět přirozený izolát a poslední kmen je konečně kýžená luminiscenční bakterie se schopností odbourávání jak naftalenu, tak salicylové kyseliny a zároveň schopná luminiscence.
U patogenních mikroorganismů je někdy určován ještě tzv. sérotyp nebo sérovar podle toho, jakými protilátkami na ně reaguje imunitní systém.
Jak vyplývá z předchozích kapitol, současná biologie zná tři domény – Bacteria, Archea a Eukarya.
Ještě nedávno se v biologii učilo (a není vyloučeno, že někde se učí dodnes), třídění života na dvě tzv. nadříše – prokaryota a eukaryota. Toto členění živých organismů už je dnes překonané. Termíny prokaryotický a eukaryotický nicméně biologie nadále používá, ale pro označování typu buňky. Bacteria a Archea mají buňky prokaryotické stavby, Eukarya mají buňky eukaryotické stavby. Stavbě buněk se věnuje detailně kapitola 2.
Hlavní rozdíly mezi doménami jsou shrnuty v následující tabulce. Některé termíny jsou vysvětleny až v dalších kapitolách a proto lze doporučit návrat k tabulce po prostudování příslušných kapitol. Tučně jsou vždy vyznačené shody mezi doménami.
|
|
Bakteria |
Archea |
Eukarya |
|
Stavba buňky |
Prokaryotická |
Prokaryotická |
Eukaryotická |
|
Operony |
Ano |
Ano |
Ne |
|
Introny |
Ne |
Ano |
Ano |
|
Vazba v membr. lipidech |
Esterová |
Éterová |
Esterová |
|
RNA polymeráza |
Jednoduchá |
Složitá |
Složitá |
|
DNA polymerázy |
Odlišné |
Podobné |
Podobné |
|
velikost malé ribozomální podjednotky |
16S |
16S |
18S |
Jak už bylo řečeno v úvodu, tradiční charakteristika organismů, které jsou zájmem objektu mikrobiologů, je velikost organismu. Tyto hranice jsou ale velmi vágní. Tradičně jsou studovány tyto skupiny organismů
Bakterie jsou jednobuněčné prokaryotické organismy s výhradně nepohlavním rozmnožováním. Jsou velmi variabilní (proměnlivé) zejména v metabolismu (pro získání energie a stavebních látek jsou schopné využít podstatně širší skupinu látek a reakcí než vyšší organismy) a fyziologii (adaptace na nejrůznější, i extrémní podmínky). V optimálních podmínkách jsou schopné daleko rychlejšího rozmnožování ve srovnání s vyššími organismy, podobně rychlejší je i jejich metabolismus.
Všechny bakterie jsou zařazeny do domény Bacteria.
Houby (fungi) jsou eukaryotické organismy mnohobuněčné či jednobuněčné, obvykle s vláknitou stavbou těla. Jejich rozmnožování může být pohlavní i nepohlavní, obvykle se obě fáze střídají. Metabolismus je poměrně uniformní, houby nejsou schopné fotosyntézy a energii získávají oxidací organických látek kyslíkem (aerobní respirací). Některé houby jsou schopné získávat energii i bez kyslíku (anaerobně) tzv. kvašením (viz kapitola 4).
Houby tvoří samostatnou říši domény Eukarya, rozdělenou na 5 oddělení (blíže viz kapitola 12). Většina hub není mikroskopická. V rámci mikrobiologie jsou tradičně zkoumány kvasinky (jednobuněčné houby obvykle s kvasnými schopnostmi) a plísně. Ty nejsou přesně vymezenou skupinou hub a jsou chápány spíše technologicky jako mikroskopické vláknité houby s negativním dopadem.
Prvoci jsou obvykle jednobuněčné eukaryotické organismy. Vývojově se jedná o samostatné menší vývojové větve. Jejich rozmnožování je obvykle nepohlavní, ve výjimečných případech i pohlavní. Jejich buňky jsou obvykle složité a mají vyvinuté speciální organely, nahrazující orgány, např. buněčná ústa či buněčnou řiť. Jejich metabolismus je obvykle založen na oxidaci potravy kyslíkem, ale známe i fotosyntetizující prvoky.
Prvoci tvoří samostatnou říši domény Eukarya, nicméně tato říše je typickým příkladem polyfyletického taxonu, protože prvoci nemají společného předka „praprvoka“. Exaktně by každá skupina prvoků měla být vlastně samostatnou říší, ale tyto říše by ve srovnání s těmi hlavními (rostliny, živočichové, houby) měly neporovnatelně méně druhů a zároveň by jich bylo příliš mnoho. Proto jsou prvoci sdruženy do jedné samostatné „nepřirozené“ říše.
Vysvětlete, které jsou tři základní domény života a jak souvisí jejich definice se stavbou a strukturou buněk.
Proč se už nevyužívá jako taxonomická charakteristika "prokaryotický" a "eukaryotický"
Definujte stručně rozdíly: bakterie x virus, kvasinka x plíseň, kvasinka x prvok
Buňka je základní stavební jednotkou života. Všechny dosud známé organismy mají buněčnou strukturu. Výjimkou jsou viry, ale u nich se vedou akademické spory o to, zda je lze vůbec považovat za živé organismy (viz kapitola 6).
Všechny známé buňky mají několik základních společných charakteristik:
Stavba a struktura buněk se liší organismus od organismu, nicméně můžeme najít některé základní společné strukturní prvky:
Všechny buňky dosud známých organismů se dají rozdělit do dvou velkých skupin – na primitivnější prokaryotické a pokročilejší a složitější eukaryotické. Hlavní rozdíly mezi prokaryotickými a eukaryotickými organismy shrnuje tabulka:
|
|
Prokaryotické buňky |
Eukaryotické buňky |
|
Vnitřní prostor |
Nedělený |
Rozdělený membránami |
|
Organely |
Nejsou |
Jsou |
|
Buněčné jádro |
Neoddělené |
Oddělené dvojitou membránou |
|
Velikost |
Menší |
Větší (řádově) |
|
Počet chromozómů |
Obvykle jeden |
Obvykle více |
|
Genetická informace |
Menší (106-107 bp) |
Větší (108-1010 bp) |
|
Plazmidy |
Časté |
Neobvyklé |
Základní chemické látky nacházené v buňkách jsou univerzální pro prokaryotické i eukaryotické buňky a pro všechny domény života.
Voda. Bez vody není život a voda tvoří dominantní součást buňky (přes 50%, ale často až 90%). Voda rozpouští většinu nízkomolekulárních látek, ve vodě jsou disociovány soli a ve vodě probíhá většina biochemických reakcí organismu.
Minerální látky. Anorganické minerální látky tvoří podstatnou část buňky. Sole jsou v buňce obvykle disociovány na kationty a anionty a tvoří základní osmotickou hladinu buňky. Mnohé kationty jsou také vázané na další organické molekuly, např. na bílkoviny. Z kationtů jsou nejzastoupenější sodný, draselný, vápenatý a hořečnatý. Z aniontů nacházíme nejvíce chloridů, fosforočnenanů (fosfátů), síranů (sulfátů) a uhličitanů.
Sacharidy. Sacharidy představují důležité energetické a stavební součásti buněk. Chemicky se jedná o hydroxyaldehydy a hydroxyketony. Podle velikosti molekul je dělíme na
Mnohé sacharidy bývají uvnitř buňky ještě všelijak modifikované, např. fosforylované, aminované apod.
Lipidy. Jako lipidy (tuky) označujeme širokou skupinu látek biologického původu, které jsou obvykle špatně rozpustné ve vodě a dobře rozpustné v jiných tucích a organických rozpouštědlech. Mezi nejdůležitější patří tzv. acylglyceroly, které jsou složeny z glycerolu, na který jsou esterovou vazbou připojeny molekuly tzv. mastných kyselin (karboxylových kyselin s dlouhým uhlíkatým řetězcem).
Aminokyseliny. Jsou jednoduché látky, v jejíchž molekulách se nachází jak kyselá karboxylová skupina ‑COOH, tak zásadití aminoskupina ‑NH2. Biologických aminokyselin je popsáno několik stovek, z nichž většina tvoří meziprodukty metabolismu. Zvláštní postavení má ale 20 aminokyselin, které tvoří základní stavební kameny bílkovin (tzv. proteinogenní aminokyseliny, také kódované aminokyseliny, protože jejich pořadí v bílkovině je zakódované v genetické informaci).
Bílkoviny (proteiny)[1]. Bílkoviny patří mezi nejdůležitější a zároveň nejvariabilnější biologické molekuly. Bez bílkovin život není, alespoň dosud neznáme žádný organismus, který by se bez bílkovin obešel. Základem struktury bílkovin je řetězec složený z 20 proteinogenních aminokyselin, které jsou vzájemně spojeny tzv. peptidovou vazbou (amidová vazba mezi karboxylovou skupinou a aminoskupinou). Pořadí aminokyselin se nazývá primární struktura bílkoviny a je až na výjimky zakódované v genetické informaci. Primární řetězec se kvůli interakcím postranních řetězců aminokyselin zkroutí do několika základních strukturních motivů, kterým říkáme sekundární struktura. Výsledný prostorový tvar molekuly se nazývá terciární strukturou. Kvartérní strukturou pak nazýváme spojení více bílkovin do jednoho funkčního celku.
Cytoplazmatická membrána představuje jakousi hranici mezi vnitrobuněčným prostorem a vnějším prostředím. Je polopropustná (semipermeabilní), tj. některé látky přes ní přejdou a jiné ne. Cytoplazmatická membrána je složená z lipidů a bílkovin, podíl bílkovin může přesáhnout i 50%.
Hlavní lipidovou složkou jsou fosolipidy.
Fosfolipidy jsou tzv. amfifilní látky, část jejich molekuly je hydrofilní (fosfát obvykle modifikovaný další polární skupinou) a část je lipofilní (zbytky mastných kyselin). Ve vodném prostředí se proto fosfolipidy samovolně organizují do tzv. fosfolipidové dvojvrstvy, ve které jsou uspořádány lipofilními konci k sobě. Tato fosfolipidová dvojvrstva je základem tzv. fluidně mozaikového modelu cytoplazmatické membrány (viz Obr. 2). Doplňují ji dále bílkoviny a u některých organismů i steroly (cholesterol, ergosterol…) a další minoritní složky. Steroly nacházíme téměř výhradně u eukaryot, prokaryota je obvykle nemají. Název fluidně mozaikového modelu vyjadřuje fakt, že uspořádání fosfolipidů a bílkovin připomíná mozaiku, celý útvar je ale při tom polotekutý (fluidní), může měnit svůj tvar a bílkoviny i fosfolipidy se neustále přemisťují.
Propustnost fosfolipidových membrán závisí na jejich
hustotě, hustší membrány jsou méně propustné. Hustotu ovlivňuje zejména míra
nasycenosti jejich mastných kyselin. Nasycené mastné kyseliny a nenasycené
mastné kyseliny s dvojnou vazbou v konfiguraci trans se v membráně lépe uspořádají a tvoří tak hustší
membránu. Nenasycené mastné kyseliny s dvojnými vazbami v konfiguraci
cis a rozvětvené mastné kyseliny mají
méně pravidelné struktury a tvoří řidší membrány. Hustota membrány může být
lokálně upravena obsahem sterolů.
Poněkud odlišnou strukturu membrán mají archea, viz kapitola 2.2.8.
Přes cytoplazmatickou membránu mohou samovolně projít pouze látky s nízkou molekulovou hmotností a bez náboje, lépe lipofilní než hydrofilní. Průchodnost závisí také na prostorové stavbě molekuly, ploché molekuly procházejí snadněji než rozvětvené. Látky procházejí dočasnými póry v membráně, popř. prostory kolem membránových bílkovin. Pro látky, které přes membránu samovolně projít nemohou, má buňka specializované bílkovinné přenašeče. Tím je zajištěno, že transportovány budou jen látky, jejichž transport buňka potřebuje.
Buněčná stěna představuje pevnou oporu buňky. Buňce také udává tvar. Pokud buněčnou stěnu opatrně odstraníme (např. účinkem některých hydrolytických enzymů), získáme tzv. protoplast, který je vždy kulatý. Kulovité uspořádání představuje energeticky nejúspornější tvar a bez tlaku buněčné stěny ho proto buňka zaujme.
Buněčná stěna dále dodává buňce mechanickou odolnost a také odolnost proti osmotickému šoku. Ocitne-li se buňka v prostředí, ve kterém je koncentrace rozpuštěných látek jiná než uvnitř buňky, dochází k samovolným procesům směřujícím k rovnovážnému stavu, tedy vyrovnání obou koncentrací. Protože ale většina rozpuštěných látek (zejména iontů) přes membránu samovolně neprochází, vyrovnávají se hladiny přesunem vody. V hypertonickém prostředí, kde je koncentrace rozpuštěných látek vyšší, pak dochází k odvodňování buňky a jejímu smrštění. V hypotonickém prostředí, kde je koncentrace rozpuštěných látek naopak nižší, dochází k přesunu vody do buňky a k jejímu bobtnání, popř. až k prasknutí. Pevná buněčná stěna takovémuto bobtnání brání a v buňce je pak permanentní osmotický tlak, který tlačí cytoplazmatickou membránu na buněčnou stěnu. Je třeba dodat, že i tato mechanická ochrana má svůj limit a v extrémně hypotonickém prostředí dochází k praskání i u buněk s pevnou stěnou. Proti hypertonickému prostředí buněčná stěna nijak nechrání a organismy žijící v takových prostředích používají jiné způsoby ochrany.
Složení buněčná stěny je různé u různých skupin organismů. Základem je ale obvykle polysacharid, často doprovázený bílkovinami nebo lipidy. Buněčná stěna rostlin je složena převážně z celulózy, buněčná stěna hub obvykle z chitinu nebo dextranů, buněčná stěna bakterií i archejí je složena z tzv. peptidoglykanu a většina živočišných buněk je obvykle bez buněčné stěny. O detailech buněčných stěn jednotlivých skupin mikroorganismů pojednávají další kapitoly.
Cytoplazma představuje polotekutou výplň buňky. Složena je zejména z vody, bílkovin, minerálních látek a meziproduktů metabolismu. V cytoplazmě probíhají mnohé biochemické reakce metabolismu.
Genetický materiál buněčných organismů je uložen v DNA (detaily viz kapitola…). Větší molekula DNA se nazývá chromozóm. Chromozómů může být v buňce i několik desítek. Většina prokaryotických buněk má jen jeden chromozóm, naopak většina eukaryotických buněk má chromozómů obvykle více, často i několik desítek. Zejména u eukaryotických buněk bývá obvyklé, že celá sada chromozómů se v buňce nachází ve více kopiích, nejčastěji ve dvou. Tento jev je nazýván ploidie Podle počtu kopií chromozómů dělíme buňky na haploidní, které mají jen jednu sadu chromozómů, diploidní, které mají dvě sady, triploidní, které mají tři sady atd. Buňky s neúplným počtem chromozómu jsou nazývány aneuploidní. Maximální dosažitelná ploidie je cca 6-8, jádrá s vyšší ploidií se rozpadají na dvě. Prokaryotické buňky jsou obvykle haploidní, eukaryotické buňky jsou buď haploidní nebo diploidní, vyšší ploidie je vzácná.
Kromě chomozómů, které představují hlavní „záznamové médium“ pro většinu genů organismu, nacházíme minoritní část genetické informace uloženou i mimo chromozómy. U prokaryotických buněk nacházíme často tzv. plazmidy, tedy krátké cyklické molekuly DNA, které kódují několik genů. Plazmidy nebo jim podobné struktury jsou u eukaryot vzácností. Mnohé organely eukaryotické buňky mají ale vlastní genetickou informaci, např. mitochondrie či chloroplasty.
Ribozómy jsou malé kuličky složené z bílkovin a ribozomální RNA (rRNA, viz kapitola…). Eukaryotické ribozómy jsou poněkud větší nicméně jejich základní struktura je stejná.
Účelem ribozómů je syntéza bílkovin podle genetické informace. Běžná buňka má řádově tisíce ribozómů umístěných volně v cytoplazmě. V eukaryotických buňkách je ale část ribozómů vázaná na membrány endoplazmatického retikula (viz kapitola….) a na těchto ribozómech probíhá syntéza membránových bílkovin.
Termínem inkluze jsou označovány malé váčky různých látek obklopené membránou. Inkluze často uchovávají zásobní živiny. Nejčastějšími zásobními látkami jsou polysacharidy, lipidy a polykyseliny.
U prokaryotických buněk nacházíme obvykle poly-b-hydroxymáselnou kyselinu (lipid), granulózu (polysacharid), glykogen (polysacharid), volutin (polyfosfát), u bakterií oxidujících síru i zásobní zrníčka síry. U eukaryotických buněk bývají zásobními látkami škrob, glykogen a triacylglyceroly.
I přes mnohé společné znaky prokaryotických a eukaryotických buněk má každý typ buněk i svá specifika, ať už v podobě speciálních struktur nebo ve velikosti.
Běžné chemické složení prokaryotických buněk je uvedeno v tabulce:
|
Látka |
Obsah |
|
Voda |
70-90% |
|
Bílkoviny |
40-80% sušiny |
|
RNA |
10-30% sušiny |
|
DNA |
2-3% sušiny |
|
Sacharidy |
5-20% sušiny |
|
Lipidy |
5-10% sušiny |
|
Popel (fosfáty, sírany, Mg, K, Na, Ca…) |
5-10% sušiny |
V porovnání s eukaryotickými buňkami (viz kapitola) mají prokaryotické buňky v sušině větší podíl nukleoých kyselin.
Tvarová variabilita není u prokaryotických buněk nijak veliká. I to je jeden z důvodů, proč se mikroskopování zejména bakterii jako přesná identifikační metody příliš nepoužívá.
Prokaryotické buňky jsou nejčastěji kulovité (koky), tyčinkovité s různou délkou, zahnuté tyčinky (vibria), spirální, mnoho buněk má nepravidelný nebo proměnlivý tvar (pleomorfní buňky). Prokaryotické buňky často zůstávají po rozdělení částečně spojené, pak vznikají nejrůznější shluky popř. vlákna.
Značná variabilita je i ve velikosti. Základní rozměry většiny prokaryotických buněk jsou v řádu několika mikrometrů (typicky 0,5x2 mm), nicméně výchylky od průměru mohou být značné. Nejmenší buňky mají rozměry v řádech desítek až stovek nanometrů, největší prokaryotické buňky jsou naopak v řádu desetin milimetru, tj. viditelné i pouhým okem.
Nukleoid představuje nepravé prokaryotické „jádro“, tedy ústřední skladiště genetické informace. Bývá obvykle přichycen k cytoplazmatické membráně a na rozdíl od pravého eukaryotického jádra není obalen žádnou membránou. Je tvořen obvykle kruhovou DNA, která je obalena polyaminy (spermin, spermidin). Polyaminy jsou zásadité, díky čemuž se dobře váží na DNA, která je kyselá díky zbytkům kyseliny fosforečné (viz kapitola…). V nukleoidu dále nalezneme malé množství bílkovin, zejména enzymů. Jelikož běžná prokaryotická DNA má délku o několik řádů (typicky 1000x) přesahující rozměry buňky, musí být důkladně sbalena a tomu právě napomáhají polyaminy a bílkoviny.
Mnoho prokaryotických buněk je pohyblivých a za tento pohyb jsou nejčastěji odpovědné bičíky (flagela, j.č. flagelum). Běžný prokaryotický bičík připomíná tvarem i funkcí lodní šroub, při pohybu se otáčí a pohání tak buňku kupředu. Bičík je ukotvený v cytoplazmatické membráně a prochází buněčnou stěnou. Bičíky jsou u bakterií složeny z bílkoviny flagelinu a bez speciálního barvení nejsou vidět v běžném světelném mikroskopu.
V jedné buňce může být mnoho bičíků, podle počtu bičíků dělíme buňky na
Některé prokaryotické buňky vytváří kolem sebe další obalovou vrstvu, slizovitý obal. Ten je složen obvykle z polysacharidů nebo bílkovin. Pro buňku znamená dodatečnou ochranu, např. před vysušením, proti chemickým látkám, usnadňuje přijímat živiny nebo funguje jako pojivový materiál, kterým se buňka přichytí např. na vhodný povrch. Pojivem jsou spojeny také buňky tvořící tzv. biofilm (viz kapitola…).
Fimbrie (též pili, j.č. pilus) jsou dlouhá dutá úzká vlákna složená z bílkoviny pilinu. Tloušťka vláken je 3-10 nm, délka až několik mm, tj. řádově srovnatelná s velikostí buňky. Funkce fimbrií není vždy zcela jasná. Někdy slouží k přichycení na povrch vhodného materiálu. Některé fimbrie (někdy nazývané „sexuální“) mají funkci při konjugaci bakterií (viz kapitola…), při které dochází k výměně genetické informace.
Mesozómy jsou vchlípeniny cytoplazmatické membrány, jejichž funkce je neznámá. Často se nacházejí blízko septa (viz kapitola…), kde se buňka začíná dělit, nebo blízko nukleoidu. Někteří vědci se domnívají, že by mesozómy mohly hrát roli v transportních procesech. Existuje ale i hypotéza, že mesozómy vznikají jen jako vedlejší produkt při barvení buněk pro mikroskopování a že tedy v živé buňce ani nejsou.
Naprostá většina bakterií má buněčnou stěnu, výjimkou jsou jen mykoplazmata, skupina primitivních parazitických bakterií. Podle typu buněčné stěny je možné rozdělit bakterie do dvou typů, na tzv. gramnegativní a grampozitivní podle toho, jak se barví tzv. gramovým barvením (viz dále). Oba typy buněčné stěny mají ale mnohé společné rysy. Gramnegativní uspořádání je evolučně starší.
Základem buněčné stěny bakterií je tzv. peptidoglykan neboli murein. Základem peptidoglykanu jsou polysacharidová vlákna složená ze střídajících se monosacharidových jednotek N-acetylglukosaminu a N-acetylmuramové kyseliny. Tato polysacharidová vlákna jsou pak prokřížena do prostorové struktury pomocí krátkých peptidů (tripeptidů až pentapeptidů). Toto prokřížení dává peptidoglykanu pevnost.
Grampozitivní bakterie mají peptidoglykanovou vrstvu velice silnou (20-80 nm), navíc vyztuženou tzv. teichoovou kyselinou. Teichoová kyselina je tvořena polyglycerolfosfátem nebo polyribitolfosfátem navázaným na aminokyseliny nebo sacharidy peptidoglykanu. Teichoová kyselina může tvořit až 50% sušiny buněčné stěny. Na povrchu peptidoglykanové vrstvy mají grampozitivní bakterie navázané další polysacharidy složené zejména z monosacharidů glukózy, manózy a galaktózy. Struktura těchto polysacharidů je specifická pro různé taxonomické skupiny grampozitivních bakterií. Tyto polysacharidy jsou také první strukturou bakteriální buňky, na kterou reaguje imunitní systém při průniku bakterie do organismu vyšších živočichů.
Gramnegativní bakterie mají ve srovnání s grampozitivními peptidoglykanovou vrstvu velice tenkou (1-3 nm) a bez teichoové kyseliny. Nad touto vrstvou mají ale ještě tzv. vnější membránu, která je složena zejména z fosfolipidů, bílkovin a lipopolysacharidů[2]. Mezi peptidoglykanovou vrstvou a vnější membránou je tzv. periplazmatický prostor, ve kterém jsou mj. umístěny některé enzymy. Tyto lipopolysacharidy představují opět první bakteriální strukturu, na kterou reaguje imunitní systém. Ve vnější membráně jsou poměrně časté póry tvořené bílkovinou porinem, které pronikají skrz periplazmatický prostor a peptidoglykanovou vrstvu až k cytoplazmatické membráně.
Barvení podle Grama je procedura, kterou objevil v roce 1884 tým dánského vědce Hanse Christiana Grama a která umožňuje rozlišit grampozitivní a gramnegativní bakterie. V době objevu nebylo známo mnoho o struktuře bakteriální stěny, nicméně Gramovo barvení se začalo používat jako jeden z diagnostických a taxonomických ukazatelů a používá se dodnes. Principem procedury je barvení usmrcené bakteriální kultury roztokem krystalové violetě a Lugolova roztoku[3] a následné odbarvení působením organického rozpouštědla, např. ethanolu nebo acetonu. Grampozitivní bakterie zůstanou filaově obarvené i po působení organického rozpouštědla, zatímco gramnegativní se odbarví. Aby byly lépe pozorovatelné pod mikroskopem i odbarvené gramnegativní bakterie, dobarví se kultura nějakým světlejším barvivem, nejčastěji safraninem na červeno.
Je třeba dodat, že příroda se ale obvykle nedá plně „zaškatulkovat“, byť se o to věda snaží. Část bakterií je proto tzv. gramlabilních a barví se podle Grama nevyzpytatelně. Svou roli hraje i stáří buněk a jejich fyziologický stav. Nejlépe se barví mladé exponenciálně rostoucí kultury. Spory grampozitivních bakterií se obvykle barví gramnegativně vzhledem k dodatečným obalovým vrstvám.
Archeální buněčná stěna není složena z peptidoglykanu jako u bakterií. Obecně známe čtyři typy archeálních buněčných stěn:
Gramovým barvením se až na výjimky většina archejí barví gram-negativně.
Archeální buňky jsou prokaryotické a jsou tak v mnoha ohledech velmi podobné bakteriálním buňkám. Kromě už diskutované buněčné stěny (viz kapitola 2.2.7.2) jsou tu ještě další rozdíly:
Základní rozdíl mezi eukaryotckými a prokaryotickými buňkami je jejich větší velikost a rozdělení vnitřního prostoru pomocí membrán na tzv. organely.
Jádro (nucleus) je organela ve které je soustředěna hlavní část genetické informace eukaryotické buňky. Je obalena dvěma membránami ve kterých se vyskytují póry sloužící zejména pro průchod mRNA ven z jádra.
Výplň jádra tvoří tzv. chromatin, což je komplex DNA a bílkovin. Hlavní podíl jaderných bílkovin tvoří histony, které se díky velkému podílu zásaditých aminokyselin (zejména lysinu a argininu) dobře váží na kyselou DNA. Dále zde nacházíme enzymy sloužící k základním genetickým pochodům (viz kapitola…). Jaderná DNA je obvykle lineární a rozdělená do více chromozómů. Vzhledem k velikosti jaderné genetické informace, přesahuje podobně jako u prokaryot délka jednotlivých molekul DNA o několik řádů velikost buňky. Pomocí histonů je proto DNA také smotaná (tzv. zkondenzovaná) do uspořádané struktury. Míra kondenzace závisí na fázi buněčného cyklu (viz kapitola…). V klidovém stavu je chromatin tzv. rozptýlený a jeho kondenzace je nižší. Při buněčném dělení tzv. spiralizuje do velmi hustě uspořádaných útvarů (chromozómů[4]), které jsou viditelné i v běžném světelném mikroskopu. Na spiralizovaném chromozómu (viz obrázek) můžeme rozeznat tzv. chromatidy a tzv. centromeru, kterou se chromozóm váže při dělení buňky na dělící vřeténko.
Mitochondrie jsou buněčné organely, jejichž hlavním účelem je zásobovat buňku energií. V mitochondriích jsou u eukaryot lokalizovány nejvýznamnější metabolické dráhy (viz kapitolu…) produkující energii, zejména buněčné dýchání, citrátový cyklus, b-oxidace mastných kyselin a produkce ATP. V běžné eukaryotické buňce jsou řádově stovky až tisíce mitochondrií podle její energetické náročnosti.
Mitochondrie je organela obalená dvěma membránami, tzv. vnější membránou a vnitřní membránou. Obě membrány mají podobnou strukturu jako cytoplazmatická membrána, základem je fosfolipidová dvojvrstva a bílkoviny. Obě membrány se ale liší propustností, zatímco vnější je velmi propustná, vnitřní membrána je extrémně nepropustná a přejdou přes ní až na výjimky jen látky, pro které existují specializované přenašeče. Vnitřní membrána je velice zprohýbaná, záhybům říkáme kristy. Mezi membránami je tzv. mezimembránový prostor důležitý pro funkci mitochondrie.
Vnitřní část mitochondrie se nazývá matrix a je tvořena hustou koloidní směsí s vysokým obsahem bílkovin (zejména enzymů).
Mitochondrie má vlastní genetickou informaci uloženou v kruhové DNA a vlastní proteosyntetický aparát. Jak DNA, tak proteosyntetický aparát je velice podobný bakteriálnímu, což podporuje endosymbiotickou teorii vzniku mitochondrií (viz kapitola 1.5 str. 4), pravděpodobnými prapředky mitochondrií jsou purpurové bakterie. Na druhou stranu mitochondrie nejsou geneticky nezávislé, část mitochodnriálních bílkovin není kódována v mitochondriální DNA, ale v jaderné DNA a do mitochondrie jsou transportovány zvenčí.
Endoplazmatické retikulum (ER) je buněčná struktura tvořená prohýbanými membránami a bílkovinami. Endoplazmatické retikulum je přisedlé k jádru a často propojuje jádro s cytoplazmatickou membránou.
Hlavním účelem endoplazmatického retikula je produkce nejrůznějších látek, které buňka potřebuje, např. membránových bílkovin, lipidů či glykogenu. Část membrán endoplazmatického retikula nese vázané ribozómy, tuto část nazýváme drsné endoplazmatické retikulum[5] a probíhá na něm syntéza membránových bílkovin. Část je bez ribozómů, nazýváme ji hladké endoplazmatické retikulum a slouží k produkci ostatních látek.
Golgiho komplex nebo též Golgiho aparát je soustava membrán a membránových váčků. Účelem kompelxu je kompletace syntetizovaných bílkovin a jejich doprava na místo určení v buňce. Golgiho komplex si vyměňuje nehotové bílkoviny s endoplazmatickým retikulem právě pomocí membránových váčků.
Vakuoly jsou buněčné vaky obalené jednou membránou. V jedné buňce může být mnoho vakuol popř. se mohou spojovat malé vakuoly do větších. Ve starých rostlinných buňkách často vyplňuje jedna vakuola prakticky celý objem buňky.
Funkce vakuol je zejména skladovací, často jsou ve vakuolách uchovány zásobní látky nejrůznějšího druhu. U rostlin mají i funkci odpadní, rostliny odpadní látky prakticky vůbec nevylučují a místo toho je hromadí právě ve vakuolách. U prvoků známe specializované vakuoly, které jsou odpovědné za buněčné trávení nebo buněčné vylučování.
Cytoskelet představuje vnitřní kostru buňky. Jedná se o soustavu vláken (filament) a trubiček (tubulů) složených z bílkovin. Jejich funkce je jednak opěrná dávající buňce tvar (což má význam zejména u živočišných buněk, které nemají buněčnou stěnu, ostatní eukaryotické buňky jsou tvarovány pevnou buněčnou stěnou). Další funkce jsou transportní, cytoskelet se např. podílí na dělení buněk posunem chromozómů děleného jádra, může způsobovat smršťování buňky apod.
Chloroplasty jsou organely tvarově podobné mitochondriím nicméně zelené barvy. Jsou obalené dvěma a někdy i třemi membránami a také mají vlastní DNA a syntézu bílkovin. Jejich účelem je fotosyntéza. Předpokládá se, že chloroplasty vznikly podobně jako mitochondrie endosymbiózou, prapředkem byly pravděpodobně sinice (cyanobakterie).
Lysozómy jsou váčky obalené membránou s bohatým enzymovým aparátem. V lysozómech probíhají zejména rozkladné procesy, např. likvidace nepotřebných bílkovin nebo rozklad fagocytovaných částic.
Většina hub jsou mnohobuněčné organismy, výjimku tvoří jen kvasinky. Houbové tělo se nazývá stélka (thalus) a jejím strukturním základem jsou vlákna (hyfy). Spleť hyf se nazývá mycelium (podhoubí). Pokud jsou hyfy nahuštěné těsně k sobě, hovoříme o sklerociu. Z mycelia někdy vyrůstají plodnice, útvary specializované na rozmnožování.
Mycelium může být v zásadě dvojího typu. Tzv. přehrádkované (septované) mycelium je tvořeno výrazně oddělenými buňkami. Nepřehrádkované mycelium naopak nemá buňky výrazně odděleny, fakticky se tedy jedná o jednu buňku s mnoha jádry.
U většiny hub jsou buňky v mycelium nejen spojené, ale zároveň i propojené pomocí pórů. Těmito póry si sousední buňky mohou vyměňovat jak látky, tak např. genetickou informaci. Takové mycelium se nazývá pravé mycelium a funguje pak jako jakési potrubí, kterým houba může transportovat živiny na velké vzdálenosti. Oproti tomu některé kvasinky tvoří útvary, které se myceliím podobají, vnitřní propojení buněk u nich ale není. Fakticky se tedy jedná o nezávislé buňky, které jsou jen spojeny popř. špatně odděleny. Takové mycelium se nazývá pseudomycelium.[6]
[1] V souvislosti s bílkovinami se používá několik pojmů. Bílkovina a protein jsou ekvivalenty a představují hotovou funkční strukturu. Peptid je označení pro několik aminokyselin spojených peptidovou vazbou, předponou může být zdůrazněno, kolik aminokyselin peptid tvoří (např. dipeptid je složen ze dvou aminokyselin, pentapeptid z pěti apod.). Polypeptid představuje více aminokyselin spojených peptidovou vazbou. Peptid je základem bílkoviny, funkční bílkovina vzniká vhodným prostorovým uspořádáním peptidů (často několika peptidů) a doplněním bílkoviny o nepeptidové složky (např. sacharidy, ionty kovů apod.)
[2] Lipopolysacharidy jsou tvořeny lipidy s navázaným polysacharidem
[3] Lugolův roztok je vodný roztok směsi jódu a iodidu draselného
[4] Termín chromozóm se u v biologii nepoužívá úplně jednoznačně. Jednak se jím nazývá každá molekula jaderné DNA. Dále se tento termín používá pro chromatin kondenzovaný do viditelných útvarů při dělení eukaryotických buněk.
[5] Názvy drsné ER a hladké ER souvisí s tím, jak se ER jeví v elektronovém mikroskopu. Membrána bez ribozómů se jeví hladká, zatímco s ribozómy se jeví zdrsněná.
[6] Pseudos znamená latinsky falešný, pseudomycelium je tedy falešné mycelium.
Definujte hlavní rozdíly mezi prokaryotickou a eukaryotickou buňkou.
Co jsou to Gram-pozitivní a Gram-negativní bakterie? Souvisí nějak tato vlastnost s taxonomickým zařazením?
Definujte hlavní odlišnosti buněčné stěny bakterií a archejí.
Vyjmenujte základní organely eukaryotické buňky a vysvětlete jejich biologický význam.
Vysvětlete hlavní rozdíly: cytoplazmatická membrána x buněčná stěna, mycelium x pseudomycelium, fimbrie x bičík
Rozmnožování patří mezi základní charakteristiky živé hmoty. Bez rozmnožování by život neexistoval. V průběhu evoluce se u organismů vyvinuly různé způsoby rozmnožování, všechny se ale dají rozdělit na dva typy – rozmnožování nepohlavní (vegetativní, asexuální) a pohlavní (generativní, sexuální).
Základní charakteristiky nepohlavního rozmnožování jsou následující:
Základními znaky pohlavního rozmnožování jsou
Pohlavní rozmnožování je evolučně mladší. Předpokládá se, že praorganismy se rozmnožovaly výhradně nepohlavně. Dnešní organismy z domén Bacteria a Archea se rozmnožují výhradně nepohlavně. Organismy z domény Eukarya se rozmnožují pohlavně i nepohlavně, oba způsoby se u nich často pravidelně či nepravidelně střídají.
Dělení buněk je základem všeho rozmnožování. U jednobuněčných organismů odpovídá buněčné dělení přímo rozmnožení celého organismu. U mnohobuněčných je předpokladem k úspěšnému rozmnožení. Buněčné dělení se významně liší u prokaryotických a eukaryotických buněk, některé zákaldní principy jsou ale totožné:
Buněčný cyklus představuje souhrn dějů vedoucí od buňky v zákaldním stavu až po dvě dceřinné buňky v základním stavu, které mohou opět vstoupit do dělení a pokračovat tak cyklicky dále. Buněčný cyklus má čtyři hlavní fáze:
Některé buňky mnohobuněčných eukaryot nejsou určeny pro další dělení (např. červené krvinky nebo povrchové buňky kůže). U těchto dále se nedělících buněk se někdy hovoří o G0 fázi.
Vlastní oddělení dceřinných buněk probíhá obvykle buď příčným dělením nebo pučením.
Při příčném dělení se buňka protáhne do délky a zhruba uprostřed se začnou objevovat vchlípeniny cytoplazmatické membrány. Tyto vchlípeniny se dále rozšiřují až dojde k oddělení cytoplazem obou buněk cytoplazmatickou membránou. Poté je dokončena buněčná stěna a buňky se mohou oddělit.
Při pučení se začne na jedné straně buňky objevovat vyboulenina (pupen). Tento pupen je postupně naplňován cytoplazmou a postupně roste. Po té, co má dostatečnou velikost, dochází k syntéze nejprve cytoplazmatické membrány a posléze buněčné stěny a buňky se mohou oddělit.
Není neobvyklé, že po oddělení cytoplazem a vzniku buněčné stěny, nedojde k úplnému oddělení buněk a ty jsou spolu dále svázány. Díky tomu vznikají nejrůznější shluky a vlákna (pseudomycelia).1.1 Růstová
křivka mikrobiální populace
Mikroorganismy jsou biologické objekty s nejrychlejším rozmnožováním, jaké známe. Mezi úplné rekordmany pak patří bakterie - některé jsou v optimálních podmínkách schopné rozdělení každou půlhodinu, tj. za hodinu se jejich počet zečtyřnásobí. Takovéto exponenciální množení samozřejmě nemůže probíhat do nekonečna, ale je limitované životním prostředím kultury a dostatkem živin.
Lag-fáze je fáze, při které se navenek neděje nic a množství organismů zůstává stejné. V této fázi se kultura adaptuje na nové prostředí (např. syntetizuje induktivní enzymy apod.) a připravuje se na dělení. Délka lag-fáze závisí na podmínkách (čím ideálnější, tím je kratší) a na velikosti inokula (větší inokulum má kratší lag-fázi).
Fáze zrychlujícího růstu. V této fázi naměříme pomalu se zrychlující přírůstek organismů. Většina organismů dokončila adaptaci a začínají se postupně dělit.
Exponenciální fáze růstu. V této fázi se organismy dělí v pravidelných krátkých intervalech (tzv. generační době) a organismů tak přibývá exponenciálně. V této fázi jsou organismy v nejlepším fyziologickém stavu a v kultuře je nejvyšší procento živých buněk. Nakreslíme-li závislost množství buněk na čase s logaritmickou časovou osou, dostaneme přímku. Proto se této růstové fázi často také říká logaritmická.
Fáze zpomalujícího se růstu. V této fázi se začíná projevovat vyčerpání živin a hromadění zplodin metabolismu, organismy se proto přestávají množit a růst se zpomaluje.
Stacionární fáze. V této fázi už nepozorujeme přírůstek organismů. Některé se sice ještě množí, jiné ale umírají a lyzují.
Fáze odumírání. V této fázi organismy už se organismům nedostává prakticky žádných živin a většina jich postupně umírá.
Vysvětlete rozdíl mezi pohlavním a nepohlavním rozmnožováním.
Které taxonomické skupiny mikroorganismů používají pohlavní nebo nepohlavní rozmnožování: bakterie, archea, houby, prvoci,viry, řasy
Vyjmenujte fáze růstové křivky a vysvětlete, v čem jsou významné z pohledu mikroorganismu-jedince a populace.
Bakterie Nitrosomonas europea má generační dobu 12 hodin. Je-li počáteční koncentrace bakterií v kalu 2.106 b/g, vypočítejte koncentraci buněk po trvání 96 hodin exponenciálního růstu. Vypočítejte specifickou růstovou rychlost bakterie.
Metabolismem je nazývána soubor všech chemických procesů, které probíhají v organismu. Na rozdíl od „běžných“ chemických reakcí mají metabolické reakce několik výrazných odlišností. Hlavním rysem metabolismu je jeho vysoká organizovanost. Metabolické dráhy jsou katalyzovány enzmy, jsou obvykle lokalizované na konkrétním místě v buňce a většina drah je nějakým způsobem regulována (řízena) na základě potřeb organismu.
Metabolismus lze rozdělit na katabolismus a anabolismus. Katabolické dráhy zahrnují rozkladné procesy, které vedou od složitějších látek k jednodušším. Jejich základní charakteristikou je konvergence, tj. z mnoha různých složitějších látek vzniká několik málo koncových produktů. Dále se obvykle jedná o reakce exergonické, tj. uvolňuje se při nich energie. Hlavním smyslem katabolismu je právě získávání energie, organismu navíc umožňuje získat některé potřebné jednodušší látky. Anabolické dráhy jsou opakem katabolických. Jedná se o dráhy syntetické. Jsou obvykle divergentní, tj. z několika málo výchozích látek je syntetizováno velké spektrum cílových látek. Anabolické reakce jsou obvykle endergonické, tj. vyžadují dodávání energie. Jejich hlavním smyslem je syntéza stavebních a funkčních látek potřebných pro život organismu.
I když je zvykem v učebnicích vykládat metabolické dráhy v podobě lineární či cyklické, ve skutečnosti jsou navzájem nejrůzněji provázané a tvoří složitou metabolickou síť.
Naprostá většina biochemických reakcí je katalyzována unikátními biologickými katalyzátory – enzymy. Ty mají proti klasickým chemickým katalyzátorům (platina, práškové železo apod.) mnoho výjimečných vlastností:
Mluvíme-li o látkách, které se účastní enzymatické reakce, hovoříme obvykle o substrátech a produktech. Substráty do reakce vstupují, produkty vystupují.
Enzymy jsou tvořeny převážně bílkovinami, známe ale i katalyticky účinné RNA, tzv. ribozymy. Enzymy se obvykle skládají z bílkovinné a nebílkovinné části. Bílkovinnou část enzymu nazýváme apoenzym, nebílkovinnou část nazýváme kofaktor. Je-li kofaktor vázán na apoenzym pevnou vazbou tak, že se od něj neodlučuje, nazývá se prostetická skupina. Je-li vázán jen přechodně po dobu reakce (čímž se prakticky chová jako další substrát), nazývá se koenzym. Na molekule enzymu nacházíme tzv. aktivní místo, ve kterém probíhá vlastní reakce. Toto aktivní místo je prostorově uzpůsobené substrátům, jejichž reakci enzym katalyzuje. Často se v tomto případě používá příměr o klíči a zámku popř. ruce a rukavici. Na mnohých enzymech nalezneme ještě další vazebná místa, např. pro látky regulující aktivitu enzymu. In vivo je běžné, že jsou enzymy navázány na membránu, buď jí prostupují (pak jsou nazývány integrálními) nebo jsou vázané na jejím povrchu (pak se nazývají periferními). Poměrně časté je také spojování několika enzymů dohromady za tvorby multienzymových komplexů.
Vzhledem k vysoké katalytické účinnosti enzymů, bývá obvykle rychlost nekatalyzované reakce ve srovnání s katalyzovanou zcela zanedbatelná. In vivo lze tak nepřesně ale prakticky správně tvrdit, že reakce bez enzymu neprobíhá.
Katalytická účinnost enzymu se vyjadřuje jako rychlost přeměny substrátu na produkty. Jednotkou je podle SI tzv. katal. Enzym má aktivitu jeden katal, pokud přemění 1 mol substrátu za jednu sekundu. Tato jednotka sice odpovídá fyzikálním standardům SI, nicméně pro enzymologickou praxi se příliš nehodí, protože je pro většinu enzymů příliš vysoká. Reálné aktivity jsou pak v řádech typu 10-15 katalů. V enzymové praxi se tak prosadila mnohořádově menší tzv. enzymová jednotka (enzyme unit) označována jako U. Enzym má aktivitu 1U pokud přemění 1 mmol substrátu za jednu minutu. Enzymové aktivity jsou obvykle udávány vztažené na množství vzorku, např. na gram či mililitr. Ještě je třeba dodat, že vzhledem k silnému vlivu teploty i dalších faktorů na aktivitu enzymů, je třeba při uvádění aktivit přidat i informaci o experimentálních podmínkách (teplota, pH apod.).
Katalytická účinnost eznymů závisí silně na vnějších podmínkách. Nejdůležitější je vliv pH, teploty a dalších látek (inhibitorů a aktivátorů). Vliv pH na aktivitu enzymů připomíná Gaussovu křivku. Existuje určité optimální pH a při jeho odchýlení aktivita přibližně rovnoměrně klesá. To je způsobeno protonací či deprotonací aminokyselinových zbytků enzymu, což má vliv na vazbu substrátu i prostorové uspořádání. Vliv teploty je podobný vlivu pH, jen pokles je po dosažení optima strmější. V tomto případě působí dva vlivy. Rychlost chemické reakce obecně roste s teplotou (což je dáno kinetickou energií molekul). Vyšší teploty nicméně způsobují denaturaci enzymu, který pak ztrácí katalytickou účinnost.
Mezi základní metabolické potřeby mikroorganismů patří
Naprostá většina mikroorganismů si dokáže svoje organické látky syntetizovat sama. Některé mikroorganismy mají ale určité anabolické dráhy zmutované a chybí jim schopnost některou látku syntetizovat. Takové organismy nazýváme auxotrofními a chybějící látku musí přijímat hotovou z prostředí. Pokud chceme zdůraznit, že určitý kmen mikroorganismů je auxotrofní, můžeme za rodovým a druhovým jménem uvést třípísmené označení chybějící látky s mínusem. Např. Escherichia coli Trp-, označuje kmen Escherichia coli, který nemá schopnost syntetizovat aminokyselinu tryptofan, je tedy auxotrof tryptofanu.
Zatímco evolučně pokročilejší organismy mají energetický metabolismus velice uniformní, u mikroorganismů nacházíme velikou rozmanitost až bizarnost. Mikroorganismy jsou schopné získat energii prakticky ze všech organických látek, ale i z mnoha anorganických. Mnoho mikroorganismů také disponuje schopností přepínat svůj základní metabolismus podle momentální situace mezi několika typy[2].
Obecně existují tři základní způsoby, kterými mohou organismy získávat energii: kvašením, dýcháním a asimilací světelné energie.
Kvašení (fermentace) je nejjednodušší způsob získávání energie. Předpokládá se, že prvotní formy života byly převážně fermentující. Principem je rozklad energeticky bohatých látek na energeticky chudší látky, přičemž energetický rozdíl organismus využije. Pro fermentaci je charakteristické, že organismus potřebuje přijímat skutečně jen jednu látku, aby získal energii. Nejčastěji jsou fermentovány sacharidy za vzniku mnoha produktů (ethanolu, mléčné kyseliny apod.), známe ale mikroorganismy schopné fermentovat např. aminokyseliny, puriny a mnoho dalších látek.
Dýchání (respirace) je proces, při kterém organismus získává energii oxidací jedné látky jinou látkou, přičemž obě získává z prostředí. Na straně redukovadla může být prakticky libovolná organická látka (sacharid, lipid, aminokyselina, organické kyseliny, aromatické i alifatické uhlovodíky, chlorované či nitrované organické látky atd.), ale i mnohé oxidovatelné anorganické látky (vodík, amoniak, sulfan, železnaté a manganaté ionty apod.). Nejúčinnějším oxidovadlem je kyslík (pak mluvíme o tzv. aerobní respiraci), ale mnohé organismy využívají i další oxidovadla, např. dusičnany, dusitany, sírany, oxid uhličitý apod. V takovém případě pak mluvíme o tzv. anaerobní respiraci.
Fotosyntéza je proces, při kterém organismy využívají energie světelných částic (fotonů). Existuje mnoho variant fotosyntézy.
Mikroorganismy jsou schopné využít řadu zdrojů uhlíku pro syntézu svých složitých organických látek. Jsou to jedna anorganické formy uhlíku (oxid uhličitý, uhličitany), tak organické formy uhlíku. Mezi nejdůležitější organické zdroje uhlíku patří sacharidy, aminokyseliny, lipidy a organické kyseliny, nicméně řada dalších organismů je schopna využít např. i uhlovodíky a jejich deriváty.
Podle složitosti využívaných zdrojů uhlíku dělíme mikroorganismy na autotrofní a heterotrofní.
Autotrfní organismy jsou takové, kterým stačí velmi jednoduché zdroje uhlíku (zejména oxid uhličitý) k tomu, aby si syntetizovaly veškeré potřebné organické látky.
Heterotrofní organismy nejsou schopné vycházet při syntézách z nejjednodušších forem uhlíku a musí proto přijímat jako zdroj uhlíku složitější organické látky. Ty pak převedou na svoje organické látky,
Metabolismus mikroorganismů je velmi rozmanitý, některé základní metabolické dráhy (byť v mírných obměnách) ale nacházíme prakticky u všech. Jedná se zejména o metabolismus sacharidů, proteinů a aminokyselin a lipidů.
Sacharidy patří k základním látkám v živých organismech. Mají jednak funkci stavební (např. v buněčných stěnách), dále zásobní (glykogen, škrob), ochranou (slizová pouzdra) a zejména energetickou.
Mikroorganismy nežijí ve vzduchoprázdnu, ale jsou v neustálém kontaktu se svým životním prostředí. Působení různých faktorů prostředí vede ke změnám ve vlastnostech mikroorganismů. Změny vlastností se dají rozdělit na dvě hlavní skupiny:
Příkladem obou typů adaptace může být adaptace na teplotu. Dojde-li např. k náhlému ochlazení, hrozí mikroorganismu ztuhnutí cytoplazmatické membrány a ztráta funkčnosti. Musí proto rychle změnit složení fosfolipidových mastných kyselin směrem k vyššímu podílu nenasycených a rozvětvených, které tuhnou při nižší teplotě. Při opětovném oteplení pak obnoví původní složení membrány. Při trvalém působení nižších teplot dochází k selekčnímu tlaku na nově vzniklé mutanty. Mutace jsou v rostoucí populaci poměrně běžné, čas od času je některá z nich pozitivní a dá vzniknout genu, který je v daném prostředí funkčnější. Tito mutanti pak mají větší konkurenční výhodu (např. se rychleji množí) a časem v populaci převládnou.
Fyziologický stav mikroorganismu koresponduje i s jeho spokojeností. Daří-li se v daných podmínkách organismu dobře, pak obvykle nespouští žádné regulační mechanismy, které by vedly ke změně. Daří-li se mu naopak špatně, snaží se regulační a obranné mechanismy aktivovat.
Termín fyziologický a nefyziologický se někdy také interpretuje jako přirozený podobně jako např. v medicíně (srovnej fyziologický – přirozený porod s nefyziologickým císařským řezem). Např. řekneme-li, že mikroorganismus je ve fyziologickém stavu, myslíme tím, že je v přirozených podmínkách a daří se mu dobře. Naopak provedeme-li nefyziologický zásah, obvykle to povede k poškození či smrti mikroorganismů.
Vnější podmínky mají vliv jak na životaschopnost organismů, tak i na jejich fyziologii. Znalost těchto vlivů je velmi důležitá pro praxi. Mnoho mikroorganismů je nežádoucích a v takovém případě je vhodné vědět, jaké podmínky jim nevyhovují (viz kapitola 9). Naopak potřebujeme-li využít mikroorganismus prakticky, např. v biotechnologických procesech, je vhodné vytvořit mu optimální prostředí. V neposlední řadě umožňuje znalost těchto vlivů odhadovat zásahy do ekosystému, kde mikroorganismy tvoří významnou součást (kapitola 10).
U každého faktoru prostředí je možné vysledovat několik stupňů působení na konkrétní organismus (viz Obr…). Organismu se nejlépe daří při optimální hodnotě faktoru a v jeho určitém okolí. Vychýlí-li se faktor příliš z normálního rozsahu, nastává stresové působení. Mikroorganismy ve stresu ještě obvykle přežívají, nicméně jejich fyziologie je narušena, např. je přerušeno dělení nebo nefungují některé metabolické dráhy. Při extrémním vychýlení působí faktor letálně a organismus zabíjí.
Teplota je dominantní faktor silně ovlivňující mikroorganismy. Závislost růstové rychlost na teplotě je obvykle nesymetrická, nižší teploty dělení pouze zpomalují, ale nebrání mu. Při extrémně nízkých teplotách už se mikroorganismus dělit nemůže, nicméně obvykle přežívá. Naopak vyšší teploty působí na mikroorganismy destruktivně, čehož se prakticky využívá při jejich likvidaci.
Z hlediska teplotní tolerance lze mikroorganismy rozdělit na několik základních skupin:
Různé skupiny mikroorganismů mají různý teplotní rozsah, ve kterém mohou přežít. Organismy vyskytující se v přírodě, mají obvykle rozsah tolerance širší a optimální teplotu spíše nižší. Oproti tomu lidské patogenní mikroorganismy jsou obvykle adaptované na přesných 37°C a i malé vychýlení této teploty vede k jejich smrti. Toho také tělo využívá zvýšenou teplotou až horečkou.
Adaptace mikroorganismů na teplotu je jednak genetická, která předurčuje mikroorganismus k životu v prostředí s určitou teplotou, a jednak fyziologická, která mu umožňuje přizpůsobit se lokálním změnám teploty. Genetická adaptace je dlouhodobá (jedná se o evoluční změnu) a zahrnuje zejména změny ve strukturách bílkovin. Bílkoviny termofilních organismů mají v molekulách více disulfidických můstků a menší podíl hydrofóbních aminokyselin. Takové bílkoviny jsou tepelně stabilnější, ale při nižších teplotách jsou rigidnější (ztuhlejší). Díky tomu se termofilní organismy při nižších teplotách nerozmnožují. Bílkoviny psychrofilních organismů jsou naopak podstatně ohebnější, v jejich molekulách je více hydrofóbních aminokyselin. Díky tomu bílkoviny při nižších teplotách neztuhnou a jsou i nadále schopné plnit svou fyziologickou úlohu. Takové bílkoviny jsou ale tepelně méně stálé.
Je třeba dodat, že pro adaptaci mikroorganismu na extrémnější teplotu je potřeba adaptace všech důležitých bílkovin zúčastněných na metabolismu. Oproti tomu mutace v některé důležité bílkovině způsobí celkovou ztrátu adaptace.
Fyziologická adaptace na teplotu zahrnuje zejména změny ve složení cytoplazmatické membrány. Ta si potřebuje uchovat svou stálou fluiditu a propustnost. Adaptace na nižší teplotu pak spočívá v náhradě nasycených a nerozvětvených mastných kyselin za více nenasycené a rozvětvené mastné kyseliny[2]. Tyto kyseliny mají nepravidelnější tvar, zabírají více prostoru a hůře se uspořádávají do krystalu. Díky tomu je jejich teplota tuhnutí nižší. Zároveň je ale takto adaptovaná membrána řidší a méně odolná proti vyšším teplotám.
[1] Termín pochází z latiny: psychros = mráz, fileo = miluji
[2] Pro praktický příklad doporučuji porovnat máslo a margarín. Zatímco chlazené máslo s převahou nasycených mastných kyselin, v lednici snadno ztvrdne, margarín z rostlinného oleje s vyšším obsahem nenasycených masných kyselin je stále dobře mazlavý.
Vysvětlete význam enzymů v biochemických reakcích.
Vysvětlete význam ATP v buněčné energetice.
Vysvětlete hlavní rozdíly: autotrof x heterotrof, respirace x fermentace.
Vysvětlete jak mikroorganismy reagují ne změnu vnějších podmínek krátkodobě a dlouhodobě.
Jaké znáte biogenní a abiogenní prvky (příklady).
[1] Chirální látka je taková látka, jejíž molekuly postrádají prvky symetrie. Molekuly takové látky pak existují ve dvou podobách nazývaných enantiomery, které jsou si vzájemně zrcadlovými obrazy. Chirální nejsou jen molekuly, ale i mnoho makroskopických objektů. Nejlepším příkladem je ruka. Ačkoliv má pravá i levá ruka stejný počet stejných prstů na stejném místě, přesto nejsou úplně stejné, jedna je prostě pravá a druhá levá.
[2] Hezkým příkladem jsou sinice. Jejich hlavním zdrojem energie je fotosyntéza produkující kyslík. Pokud ale není dostatek světla, mohou přepnout metabolismus na oxidační či fermentační.
Přenos genetické informace z generace na generaci (dědičnost) je jedním ze základních atributů živé hmoty. Způsob, jakým je genetická informace v organismech ukládána a předávána dalším generacím, je u všech organismů velice podobný (liší se jen detaily) a svědčí o stejném původu všech organismů
Genetická informace všech organismů je uložena v tzv. nukleových kyselinách. Jejich název je odvozen o latinského nucleus, což znamená jádro. Označení vychází z toho, že většina genetické informace je uložena právě v jádře buňky.
Nukleové kyseliny jsou makromolekuly s velmi rozdílnou velikostí a s pravidelně se opakující strukturou. Základem struktury je tzv. pentóza-fosfátový skelet, tedy pravidelné střídání zbytku kyseliny fosforečné a sacharidu. Na sacharidu je pak glykozidickou vazbou navázána tzv. báze, což je derivát pyridinu nebo purinu. Pořadí různých bází pak určuje vlastní genetickou informaci.
V živých organismech se vyskytují dvě základní nukleové kyseliny – deoxyribonukleová kyselina (DNA – deoxyribonucleic acid) a ribonukleová kyselina (RNA - ribonucleic acid). Jejich základní odlišnost je v použitém sacharidu a v uplatňujících se bázích.Viry jsou nebuněčné částice, o jejichž zařazení do biologického systému se stále diskutuje. Většina vědců se dnes přiklání k tomu, že viry nelze označit za živé organismy, protože nesplňují základní atribut živé hmoty a to schopnost samostatného rozmnožování. Hlavním důvodem je absence proteosyntetického aparátu. Viry proto parazitují v živých buňkách a využívají jejich proteosyntetický aparát. Taková činnost buňce obvykle škodí.
Název virus pochází z latiny a znamená jed. První virus popsal už v roce 1892 ruský vědec Dimitrij Ivanovský. Ten zkoumal chorobu tabáku (tzv. tabákovou mozaiku) a zjistil, že i když z tabákové šťávy odstranil bakterie filtrací přes porcelánový filtr, stále byla tato šťáva infekční. Choroba tedy byla způsobena infekční částicí menší než je bakterie[1].
Prakticky libovolné buňky mohou být v principu hostiteli virů. Viry jsou nicméně selektivní k hostitelským buňkám, které infikují, tj. určitý typ viru napadá jen určitý typ buněk. Selektivita je dána tím, že virus se musí při průniku do buňky navázat na určitou povrchovou strukturu buňky. Tato struktura se nazývá receptor a virus je obvykle schopen rozpoznat jen jeden receptor. Nejlépe prostudované jsou viry živočichů (zejména lidské), rostlin a bakterií (bakteriofágy), poněkud méně informací je o virech hub (mykofágy) a archejí, nicméně i tyto byly popsány.
O evolučním původu virů není dnes prakticky nic známo, pravděpodobně se jedná o artefakty buněk, schopné parazitického života. Viry mají naopak vliv na průběh evoluce, v genomu většiny organismů nacházíme typické virové sekvence. Podle některých teorií se viry mohly podílet na tzv. horizontálním přenosu genů mezi různými organismy, mohly způsobovat mutace, mohly přispívat k rozšíření genetické informace apod.
Viry jako buněční parazité cílovým buňkám obvykle škodí, často je ničí. Není proto divu, že spousta virů způsobuje více či méně závažné choroby jak živočichů včetně člověka, tak i rostlin a hub. Mezi tyto choroby patří např. různá nachlazení, chřipka, hepatitida (žloutenka), AIDS, vzteklina atd. Některé viry mohou způsobovat i rakovinu díky zásahu do genomu hostitelské buňky.
Viry jsou rozmanitých velikostí, od cca 20 nm, které je možné vidět pouze elektronovým mikroskopem, až po 800 nm, které lze vidět i v kvalitním světelném mikroskopu.
Konkrétní virovou částici nazýváme virion. Každý virion je složen z jedné nebo několika molekul nukleové kyseliny, která nese jeho genetickou informaci. Tato nukleová kyselina je pak obvykle obalena bílkovinným obalem, tzv. kapsidou. Tvar kapsid je rozmanitý, nejčastěji nacházíme dvacetistěn (eikosaedr), ale známe i viry kulaté, spirálovité či ve tvaru střely.
Kromě bílkovin kapsidy bývají součástí viru i další bílkoviny, sloužící zejména k infekci hostitelské buňky. Některé viry bývají ještě obaleny fosfolipidovou membránou, která pochází z hostitelské buňky. Takové viry nazýváme obalené.
Zatímco všechny známé buněčné organismy mají genetickou informaci uložené téměř výhradně v dsDNA, u virů nacházíme daleko větší rozmanitost. Viry uchovávají svou informaci jak v DNA, tak v RNA, přičemž obě mohou být jak dvouvláknové, tak i jednovláknové. U vláken RNA pak ještě rozlišujeme tzv. +RNA (pozitivní), která může sloužit přímo jako mRNA pro syntézu bílkovin, a tzv. --RNA (negativní[2]), jejíž informace musí být nejprve přepsána do mRNA, aby podle ní mohly být syntetizovány bílkoviny.
Genetická informace virů zahrnuje obvykle jen několik málo genů. Mezi nimi nacházíme bílkoviny kapsidy, dále bílkoviny odpovědné za rozpoznání hostitelské buňky, a občas i některé enzymy, např. speciální transkriptázy a polymerázy. U virů nikdy nenacházíme žádné části translačního aparátu (geny pro tRNA, rRNA a ribozomální proteiny). V syntéze bílkovin jsou tedy viry přímo závislé na hostitelské buňce.
Některé viry mají svou genetickou informaci zakódovanou ve více molekulách nukleových kyselin. Takový genom pak nazýváme segmentovaný. Příkladem je např. chřipkový virus, který má sedm segmentů -RNA, z nichž každý nese 2-3 geny.
U virů nacházíme obvykle velice zhuštěný zápis genetické informace do nukleových kyselin. Není tak neobvyklé, že na dsDNA/dsRNA jsou exprimována ve stejném místě obě vlákna, přičemž každé dává vzniknout jinému proteinu s jinou funkcí(!). Takováto nukleová kyselina je pak nazývána dvojsmyslná. Není také neobvyklý překryv sousedních genů, kdy produkované bílkoviny mají společnou část primární struktury. U některých virů (např. HIV) nacházíme dokonce překryv genů způsobený posunem čtecího rámce. Začátek čtení pak rozhoduje, jaký protein vznikne.
RNA je méně stabilní molekula ve srovnání s DNA a tak viry, které používají RNA jako nositelku genetické informace, podléhají často mutacím. To způsobuje, že se objevují stále nové a nové varianty viru, které mohou způsobit nové epidemie. Exemplárním příkladem je opět chřipkový virus, jehož nová varianta se objevuje cca 1-2x do roka. Očkování proti chřipce proto není trvalé a musí se pravidelně obnovovat. Variabilitě chřipkových virů navíc přidává segmentování jejich genomu, které umožňuje kombinovat segmenty z různých virů tehdy, je-li jedna buňka současně infikovaná dvěma různými chřipkovými viry. Časté mutace RNA virů také způsobují to, že lépe unikají imunitnímu systému. Některé virové infekce pak přecházejí do chronického stádia (např. některé hepatitidy).
Viry jsou velice rozmanité a jejich životní cykly jsou proto velice pestré. V této pestrosti ale můžeme vysledovat dvě základní životní schémata . Tzv. lytické infekce končí lýzí[3] a smrtí buňky, zatímco tzv. latentní infekce (též nazývaná mírná a takový virus mírný) smrtí buňky nekončí.
Typická lytická infekce probíhá obvykle následujícím způsobem:
Mírná infekce může probíhat podobně až do fáze 4. nebo 5. Produkce nových virionů může být natolik pomalá, že jsou uvolňovány do prostředí průběžně a buňka nelyzuje. Často také dochází k tomu, že virová nukleová kyselina zůstává v buňce bez exprese (takovou genetickou informaci nazýváme provirus). Virová nukleová kyselina se může buď integrovat (začlenit) do DNA buňky a stane se tak její součástí, popř. může zůstávat bez integrace v buňce jako nezávislá molekula DNA/RNA. „Spící“ provirus se může za určitých okolností „probudit“, např. mutací, či fyziologickou změnou v buňce (stres apod.).
Integrace viru do hostitelské DNA může u vyšších organismů způsobit také rakovinu. Stane se tak tehdy, pokud vložená virová DNA přeruší některý z tzv. onkogenů, tedy genů sloužících k regulaci buněčného dělení. Je-li poškození velké (např. při současném poškození několika onkogenů i jiným způsobem, např. v důsledku kouření[4]), pak dojde k nekontrolovanému buněčnému bujení – rakovině.
Virus integrovaný do buněčné DNA je pochopitelně předáván dalším generacím buněk. DNA vyšších organismů obsahuje virových sekvencí spoustu. Předpokládá se, že viry mají význam v evoluci kvůli tzv. horizontálnímu přenosu genetické informace[5]. K němu může dojít např. chybou při tvorbě nových virionů, kdy se do viru dostane i část „normální“ DNA buňky a je pak integrována do dalšího hostitelského organismu. I když je takováto událost málo pravděpodobná, tak při běžných virových infekcích vznikají stovky miliónů nových virionů a evoluce měla na podobné přenosy času také dost.
Vzhledem k tomu, že viry nemají proteosyntetický aparát a tudíž ani ribozomální RNA, nelze je zařadit do univerzálního fylogenetického stromu (viz kapitola 1.4), který je právě na sekvencích rRNA založen. O evoluci virů se navíc neví prakticky nic. Taxonomický systém virů je proto umělý založený zejména na
Bakteriální viry patří mezi poměrně dobře prozkoumanou skupinu virů. V některých případech se staly dokonce modelovými organismy pro výzkum. Hlavní výhoda je ta, že tyto viry napadají pouze bakterie a nehrozí proto riziko nákazy pro výzkumné pracovníky. Mnohé viry mají poměrně široké spektrum svých hostitelů, jiné zase úzké. Ekologická role těchto virů není příliš prozkoumána a spočívá mj. v udržování rovnováhy v bakteriálních společenstvech. Bakteriofágy jsou někdy používány v medicíně proti patogenním bakteriím.
Archeální viry jsou popsané, ale nepříliš prozkoumané, což souvisí s obecně menším množstvím informací o archejích obecně.
Viry rostlin bývají obvykle neobalené, protože obalení obvykle překáží pevná celulózová stěna rostlinných buněk. Jejich činnost je zejména negativní, způsobují nejrůznější choroby rostlin a tím např. snižují výnosy zemědělských plodin.
Virus tabákové mozaiky (tobamavirus) je prvním objeveným virem a zároveň je také mezi rostlinnými viry nejprozkoumanější a slouží jako modelový objekt při výzkumu. Má mezi viry jedny z největších virionů. Kapsida je helikální (spirálovitá) a obklopuje ssRNA.
Obr.1. Virus tabákové mozaiky (Tobamavirus)
Viry živočichů bývají častěji obalené, protože živočišné buňky nemají buněčnou stěnu a obalení viru zbytky membrány tak nic nebrání. Vzhledem k jejich významu v humánní i veterinární medicíně patří mezi hodně zkoumané objekty. Tradičně jsou děleny ještě na viry obratlovců a viry bezobratlých.
Papilomavirus je neobalený virus, jehož genetickou informaci tvoří dsDNA. U lidí způsobuje např. bradavice, nicméně je to zároveň onkogenní virus, který může způsobit rakovinu děložního čípku.
Poliovirus je neobalený virus s genetickou informací v ssRNA. Způsobuje např. dětskou obrnu.
Herpesvirus je obalený virus s genetickou informací v dsDNA. Tato DNA může zůstávat v buňce v latentním stavu a aktivovat se jen občas. Herpesviry způsobují např. plané neštovice a opary (jak klasický opar na rtech, tak nebezpečnější pásový opar). Promořenost lidské populace herpesviry je značná.
Orthomyxovity (viry chřipky) jsou obalené viry s genetickou informací v segmentované ssRNA. Existuje ptačí a savčí forma tohoto viru, některá zvířata jsou ale vnímavá k oběma typům virů. Virus značně mutuje a objevují se různě nebezpečné formy toho viru, některé téměř neškodné, jiné naopak s velkou úmrtností.
Retroviry jsou skupinou obalených virů, jejichž genetická informace je uložena v ssRNA. K její expresi využívá virus neobvyklý způsob, tzv. reverzní transkripci katalyzovanou enzymem reverzní transkriptázou. Při tomto procesu dojde k přepisu virové ssRNA do DNA a následné integraci na náhodné místo hostitelské DNA. Retroviry proto patří mezi onkogenní viry, které mohou způsobit rakovinu. Nejznámějším retrovirem je HIV (Human Immunodeficiency Virus), způsobující obávanou chorobu AIDS. Reverzní transkriptáza retrovitů je poměrně nepřesný enzym, který navíc nekontroluje chyby při přepisu. Díky tomu vznikají stále nové a nové varianty viru a ten tak může unikat imunitnímu systému. HIV infikuje T-lymfocyty a vyřazuje je tak z normální činnosti proti vetřelcům. Spolu se vznikem nových variant virů to vede k postupné kolonizaci a likvidaci celého imunitního systému. Nemocný AIDS pak obvykle umírá na nějakou banální nemoc typu rýma.
Definujte základní rozdíly mezi DNA a RNA.
Struktura DNA a RNA je z chemického pohledu velice podobná. Zamyslete se, proč je RNA daleko méně stabilnější než DNA (nápověda: souvisí to s biologickou funkcí obou nukleových kyselin).
Vysvětlete smysl a biologický význam základních genetických pochodů (transkripce, translace, replikace).
Vysvětlete, proč jsou viry označovány za buněčné parazity. Proč někteří vědci nepovažují viry za živé organismy?
[1] I když Ivanovského objev byl víceméně náhodný, není úplně náhoda, že byl jako první objeven právě virus tabákové mozaiky (Tobamavirus). Je to totiž jeden z největších známých virů a na rozdíl od jiných virů je vidět v kvalitním světelném mikroskopu.
[2] Pro lepší pamatování doporučuji mnemotechnickou pomůcku – fotografii. Podobně jako pozitiv představuje skutečný obraz, tak i pozitivní RNA může sloužit přímo pro syntézu bílkovin. Oproti tomu fotografický negativ obsahuje převrácenou informaci o barvách, která se převrací zpět do původních barev v procesu vytváření fotek. Podobně negativní RNA je třeba nejprve přepsat do komplementární („převrácené“) mRNA a pak je teprve možné ji překládat do bílkovin.
[3] Lýze buňky znamená porušení jejich obalů (cytoplazmatické membrány a buněčné stěny) a následné vylití obsahu cytoplazmy do prostředí. To vede nejčastěji ke smrti buňky.
[4] Vznik rakoviny se dá přirovnat ke kurzovému sázení, i když s opačným výsledkem. Sázkař potřebuje k výhře správně tipnout všechny výsledky na tiketu a i jediná chyba mu výhru zhatí. Podobně ke vzniku rakoviny je potřeba několik současných nešťastných událostí v buňce, např. několik současných mutací onkogenů a zároveň oslabení imunitního systému, který včas rakovinou buňku nerozpozná a nezlikviduje. Pravděpodobnost („kurz zápasu“) mnohých těchto událostí lze ovlivňovat chováním, např. omezením přísunu rakovinotvorných látek v důsledku kouření, správnou životosprávou apod.
[5] Horizontální přenos genů znamená přenos mezi nepříbuznými jedinci. Oproti tomu vertikální přenos genetické informace je přenos na potomky.
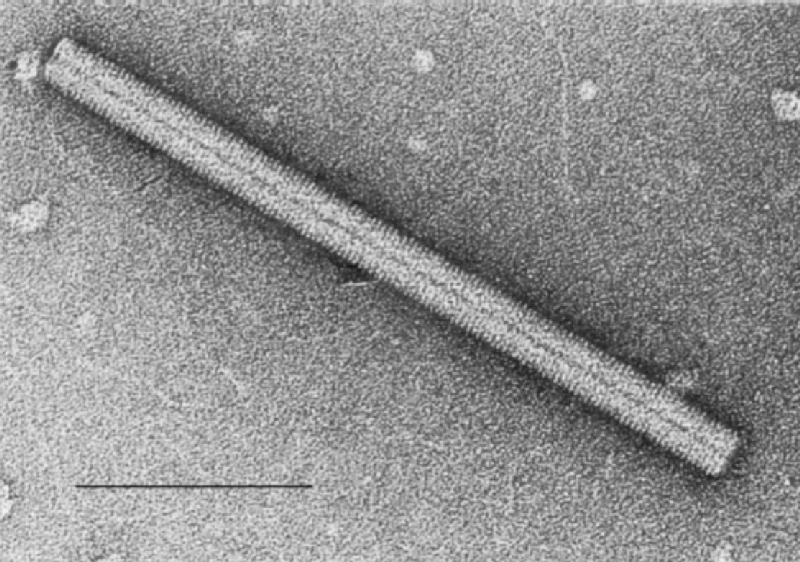 Virus tabákové mozaiky (Tobama virus)
Virus tabákové mozaiky (Tobama virus)Mikrobní ekologie je hraniční obor na pomezí mikrobiologie, ekologie a dalších biologických disciplín. Hlavní náplní je zkoumání vzájemných vztahů
Populace je skupina jedinců jednoho druhu obývajících určité území.
Společenstvo je soubor druhů, které se vyskytují ve stejném čase na stejném území. Společenstvo je tvořeno populacemi.
Mikroorganismy nacházíme prakticky všude a není tak divu, že běžně tvoří nejrůznější společenstva. Ty je ale třeba studovat z většího nadhledu (ne jen mikrobiologického), protože jejich součástí bývají i vyšší eukaryotické organismy (houby, rostliny či hmyz) a bez komplexního studia není možné správnou funkci společenstva pochopit.
Mezi členy společenstva je možné nalézt prakticky všechny možné varianty vzájemných vztahů, od vzájemně kooperace až po vzájemné škození. Tyto vzájemné vztahy shrnuje tabulka:
|
|
+ |
0 |
- |
|
+ |
Mutualismus Symbióza |
|
Parazitismus |
|
0 |
Synergie Syntrofismus |
Neutralismus Komenzialismus |
|
|
- |
|
|
Antagonismus |
Neutralismus a komenzialismus znamená vzájemné neovlivňování populací.
Synergií se rozumí stav, kdy činnost jedné populace prospívá jiné populaci bez vlastního užitku. Příkladem může být zvýšení teploty prostředí intenzivním metabolismem (např. v kompostu), což zlepšuje životní podmínky pro přítomné termofilní (teplomilné) populace.
Syntrofismus je synegie uplatňovaná v potravním řetězci. Jedná se zejména o stav, kdy jedna populace využívá odpadních produktů metabolismu jiné populace. Příkladem může být methanogeneze, kdy methanogenní archea využívají odpadní oxid uhličitý a vodík produkovaný jako konečný produkt fermentačního metabolismu některými rody bakterií Clostridium.
Mutualismus znamená vzájemné pozitivní ovlivňování populací.
Symbióza je vyšší forma mutualismu, při které dochází k výhodné spolupráci dvou jedinců různého druhu nebo dvou populací. Často bývá alespoň pro jednoho symbionta nepostradatelná popř. výrazně zvyšuje jeho životaschopnost. Některé příklady jsou uvedeny v dalších kapitolách.
Parazitismus znamená využívání látek jednoho organismu (hostitele) druhým organismem (parazitem). Hostiteli činnost parazita škodí. Rozlišujeme endoparazity, kteří žijí v těle hostitele, a ektoparazity, kteří žijí na hostitelově povrchu.
Predace je stav, kdy je jeden organismus (kořist) zabit a zkonzumován druhým organismem (predátorem). Mezi mikroorganismy moc predátorů nenacházíme, zato role kořisti bývá poměrně častá.
Antagonismus znamená vzájemné negativní ovlivňování populací. Příkladem je např. antagonismus bakterií a saprotrofních hub při kolonizaci opadu (např. spadaného listí). Houba produkuje extracelulární enzymy rozkládající polysacharidy na oligosacharidy až monnosacharidy, které pak konzumuje. Bakterie tyto monosacharidy houbě fakticky kradou, čímž jí ochuzují o získané živiny. Mnoho hub proto produkuje do prostředí i antibiotika, která činnost bakterií tlumí.
Příroda je velice variabilní a nabízí mnohem více vzájemných vztahů, než kolik se dá zařadit pod příslušné termíny. Určení přesného typu vzájemného vztahu je proto často obtížné až nemožné. U některých vztahů je navíc hranice mezi prospěchem a škozením velice vágní. Jeden organismus může druhému jak škodit tak prospívat, přičemž určení převládajícího směru je značně obtížné.
V reálném životním prostředí panuje velká konkurence mezi organismy, mikroorganismy nevyjímaje. Hlavní „boje“ se vedou o živiny, vodu a životní prostor. Strategií těchto konkurenčních bojů je nepřeberné množství a jejich výzkum přináší pravidelná zajímavá odhalení. Mezi nejobvyklejší strategie mikroorganismů patří
Při zkoumání intenzity rozmnožování, kvality potomstva a rychlosti osidlování nových lokalit lze rozlišit tři základní typy růstových strategií.
K-stratégové mají obvykle pomalejší rozmnožování, tedy i méně vyprodukovaného potomstva za jednotku času, nicméně kvalita tohoto potomstva bývá velká a velká část potomků má šanci na přežití. K-stratégové nemají obvykle velkou tendenci migrovat. Při kolonizaci nového prostředí tvoří zprvu minoritní složku, která ale časem (po uchycení dostatečně velké populace) může získat na dominanci.
R-stratégové jsou opakem k-stratégů. Rozmnožují se obvykle velice rychle, tvoří velké množství potomstva jehož kvalita ale není velká a velká část potomstva se nedožije dospělosti. Tyto organismy dávají obvykle velký důraz na mobilitu a velké množství potomstva jim umožňuje rychle kolonizovat nová prostředí. V těch zaujmou zpočátku dominantní postavení, které ale časem obvykle ztrácejí.
S-stratégové osidlují obvykle takové lokality, ve kterých jsou životní podmínky značně stresové. Adaptace těchto mikroorganismů je obvykle danému prostředí přizpůsobena, zatímco konkurence ne. Konkurentů je tedy méně.
Je třeba zdůraznit, že přesné vymezení organismu jako K- či R-stratéga není obvykle jednoznačné. Nejvhodnější je porovnat dva organismy a určit růstovou strategii relativně. Taková houba bude např. ve srovnání s lesní faunou typický R-stratég, protože chrlí do prostředí milióny spór, z nichž se málokterá uchytí. Ve srovnání s bakteriemi je ale spíše K-stratég. protože bakterie se rozmnožují daleko rychleji. Uchycené houbové mycelium pak ale vydrží daleko nepříznivější podmínky,
Pohyb není u mikroorganismů nijak významným faktorem. Většina bakterií stráví celý svůj život na malém území. Ani pohyblivé bakterie obvykle necestují na velká vzdálenosti. Za mikrobní pohyb lze považovat i pasivní přesun, např. spór větrem či v těle nebo na těle nějakého vyššího organismu. Za formu transportu lze považovat i přesun živin houbovými mycelii na velké vzdálenosti.
Monopol je dobrý způsob, kterak nebojovat s konkurencí. Mezi mikroorganismy nacházíme mnoho skupin, které jsou schopné využívat živiny, které jiné organismy využít nedokáží nebo jim dokonce škodí. Mnohé bakterie umí např. rozkládat uhlovodíky a mnohé další organické látky, které jsou pro jiné organismy (zejména pro vyšší organismy) toxické. Do této kategorie lze zařadit např. jedinečnou schopnost některých hub rozkládat lignocelulózu (tj. hlavní složku dřeva) nebo fotosyntézu, která otvírá cestu k téměř nevyčerpatelnému zdroji energie. Týká se i dalších živin, např. schopnost asimilace anorganických zdrojů uhlíku a síry rostlinami a mikroorganismy je staví před konkurenci živočichů (které jsou schopné využít pouze organicky vázaný dusík a síru). Některé plísně jsou zase schopné využívat vzdušnou vlhkost a mohou tak růst na relativně suchých substrátech.
Negativní ovlivňování jiných jedinců a populací je v přírodě velice rozšířené. Mikroorganismy obvykle do prostředí produkují inhibující látky (antibiotika, toxiny nebo některé rozkladné enzymy), které jsou pro jiné populace toxické. U některých hub nacházíme mechanismus inhibice růstu hyf přímým dotykem hyfy konkurenční houby. Do této kategorie patří i aktivita mnohých mikroorganismů vedoucích k úpravě životního prostředí na takové, které jim lépe vyhovuje. Např. bakterie rodu Thiobacillus produkují do prostředí kyselinu sírovou, čímž vytvářejí silně kyselé prostředí, na které jsou dobře adaptovány a které většinu konkurenčních mikroorganismů ničí.
Popisem nejroztodivnějších forem spolupráce mezi organismy by se daly popsat tuny papíru. Mezi mikroorganismy jsou velice rozšířené zejména potravní řetězce (syntrofie), kdy organismy v řetězci postupně využívají svých odpadních produktů. Při rozkladu komplezních substrátů se uplatňují různí členové společenstva, izolované populace by tento substrát obvykle nedokázaly sami rozložit. O některých zajímavých symbiózách pojednávají další kapitoly.
Ochrana vlastního teritoria je záležitost známá spíše u vyšších organismů, zejména živočichů. Různé varianty ale nacházíme i u mikroorganismů. Mnoho bakterií je schopno vytvářené tzv. biofilmy, tj. husté a velice odolné vrstvičky buněk a slizu, kterými jsou schopné obalit „svůj“ substrát a zabránit tak dalším mikroorganismům v přístupu k němu. Mnohé houby podobně obalují „svůj“ substrát myceliem, které navíc obvykle produkuje i nějaké inhibující antibiotika. U kvasinek v ovoci bohatém na sacharidy dochází často ke kvašení i v přítomnosti kyslíku (tzv. Crabtreeho efekt, viz kapitola…). Předpokládá se, že vyprodukovaný ethanol brání dalším mikroorganismům ve využití tohoto ovoce, zatímco kvasinky jsou proti ethanolu odolnější.
Půda představuje svrchní část zemské kůry. Je to hmota složená zejména ze zvětralých hornin, vody, vzduchu a dalších organických i anorganických látek obývaná společenstvem organismů. Vlivem vnějších podmínek a činností organismů prochází půda neustálými přeměnami.
Půda je vertikálně rozdělena na vrstvy, tzv. horizonty. Různé půdy obsahují různé horizonty, obecně nacházíme tyto (označují se mj. jednopísmennými zkratkami):
U neoraných (zejména lesních) půd lze v organickém horizontu obvykle rozeznat ještě další podvrstvy:
Mikroorganismy tvoří dominantní část živých organismů v půdě. V jednom gramu půdy nacházíme typicky 108-109 bakterií, 105-106 hub, 104-105 prvoků a 104-105 eukaryotických řas.
Množství mikroorganismů, jejich druhová skladba, bohatost společenstva i jeho aktivita jsou značně proměnlivé u různých půd. Obecně lze ale vysledovat tyto zákonitosti:
Mikroorganismy plní v půdě různé funkce v rámci půdních společenstev, mezi nejdůležitější )ale ne zdaleka všechny) patří tyto:
Půdní mikroorganismy se mohou nacházet v různě aktivním stavu od vysoké aktivity (např. na podzim po spadu čerstvého listí) až po téměř nulovou aktivitu (např. v zimě nebo za vysokého sucha). Mikrobiální aktivita je ovlivněna mnoha faktory, mezi nejdůležitější patří:
Rozklad organické hmoty je stěžejní ekologická role mikroorganismů. Zástupci nejrůznějších skupin mikroorganismů (bakterie, houby…) jsou schopné rozkladu velkého množství organických látek z nichž mnohé nejsou odbouratelné jinými organismy.
Rozkládané látky je možné rozdělit na přírodní a vyprodukované člověkem (anthropogenní). Ty bývají také nazývány polutanty (z latinského polluo = znečistit).
Dřevo je jeden z nejrozšířenějších přírodních materiálů na zemi. Jeho rozklad ale není nijak jednoduchý.
Dřevo se skládá ze tří hlavních složek – celulózy (40-50%), ligninu (20-30%) a hemicelulóz (20-30%) a mnoha minoritních látek, organických (1-3%), minerálních (0,1-0,5%). Dřeva různých stromů se liší poměrem hlavních složek, např. tropická dřeva mají obvykle mnohem více organických látek než dřeva stromů mírného pásu.
Celulóza (poly-b-1,4-glukopyranóza) je krystalický polysacharid složený z glukózových jednotek spojených b-1,4-glykosidickou vazbou. Běžná molekula celulózy má relativní molekulovou hmotnost 300 000 – 500 000, její řetězce nejsou rozvětvené a je nerozpustná ve vodě.
Hemicelulózy jsou polysacharidy o řádově nižší molekulové hmotnosti než celulóza s variabilnější strukturou. Molekuly hemicelulóz jsou lineární i rozvětvené. Převládající monosacharidy jsou D-xylóza, glukóza a manóza, ale nacházíme i minoritní složky, např. arabinózu, galaktózu nebo kyselé sacharidy. Nejběžnějšími hemicelulózami jsou xylany (poly-D-xylóza) a glukomanany (kopolymer D-glukózy a D-manózy.
Lignin je nepravidelný kovalentní polymer, jehož dominantní složkou jsou fenoly různě substutuované zejména alkylovými a alkoxylovými skupinami. Lignin je tmavé až černé barvy a je nerozpustný ve vodě.
Lignin a celulóza tvoří ve dřevě obtížně oddělitelný tzv. lignocelulózový komplex, někdy nazývaný
jen lignocelulóza. Celulóza udává
dřevu pevnost, lignin ovlivňuje tvrdost a dodává dřevu hydrofobicitu
(vodoodpudivost) a hemicelulózy spoluovlivňují tvrdost dřeva.
Na rozkladu dřeva se podílejí bakterie a nejvýraznější mírou houby. Některé bakterie disponují celulolytickými enzymy, které mohou rozkládat celulózu, zejména tu, která není příliš vázána na lignin. Mnohé bakterie jsou také schopné rozkládat hemicelulózy. Není dosud známa bakterie schopná kompletního rozkladu dřeva bez spoluúčasti jiných mikroorganismů.
V říši houby najdeme skupiny, které jsou schopné daleko účinnějšího rozkladu dřeva popř. dokonce kompletního rozkladu dřeva. Tzv. dřevokazné houby, tedy ty se schopností rozkládat dřevo alespoň částečně. Podle rozkládaných složek dělíme dřevokazné houby na několik skupin:
Odbourávání dřeva je aerobní proces, který probíhá zejména působením extracelulárních enzymů. Rozkládající organismy je produkují do prostředí, kde působí rozkladné procesy. Rozložené nízkomolekulární komponenty jsou pak organismem absorbovány a slouží jako výživa.
Odbourávání ligninu je přísně aerobní proces, při kterém hrají hlavní roli oxidační enzymy dřevokazných hub, zejména oxidázy a peroxidázy. Jejich specifita bývá velice nízká a rozkládají nejrůznější chemické struktury. Peroxid vodíku vzniká jako produkt oxidačních reakcí za účasti kyslíku. Mezi nejdůležitější enzymy patří:
Odbouráváním ligninu vznikají tzv. huminové látky (viz dále) s nižší molekulovou hmotností než měl výchozí lignin. Houba nezískává rozkladem ligninu žádnou energii a prakticky ani jiné živiny (výjimkou jsou některé dusíkaté látky vázané v ligninu, nicméně se jedná o minoritní zdroj), naopak musí na štěpení energii vydávat. Hlavním důvodem rozkladu ligninu je obnažení celulózy, kterou pak houba rozkládá a využívá jako zdroj uhlíku a energie.
Odbourávání celulózy je doménou bakterií a hub. Štěpení probíhá pomocí extracelulárních enzymů, tzv. celuláz:
Některé houby jsou schopné alternativního neenzymatického štěpení celulózy radikálovými reakcemi za účasti peroxidu vodíku a železnatých iontů. Toto štěpení je značně nespecifické a štěpí i hemicelulózy a další složky dřeva.
Odbourávání hemicelulóz je záležitost mnoha extracelulárních enzymů. Mezi nejdůležitější patří:
U těchto enzymů dochází k častému překrývání aktivit (např. glukosidáza je schopna s nižší rychlostí odštěpovat i galaktózu) a jsou velice variabilní mezi jednotlivými rody mikroorganismů.
Opadem se myslí odumřelé části zejména rostlin, jako např. spadané listí, větvičky, uhynulé rostliny a další materiál, který už nelze označit za plně živý. Rozklad opadu stojí na začátku důležitého potravního řetězce, který je nazýván detritový (z latinského detritus = drť). Organismy rozkládající opad se nazývají saprotrofní nebo saprofytické a mají zástupce mezi bakteriemi, houbami, prvoky i vyššími živočichy. U mikroorganismů probíhá rozklad opadu opět pomocí extracelulárních enzymů, přičemž enzymová výbava různých saprotrofních mikroorganismů je značně variabilní. Protože součástí opadu bývá i dřevo resp. dřevu podobné struktury, uplatňují se mj. i lignolytické a celulolytické enzymy.
Složení běžného opadu je uvedeno v tabulce:
|
Složka |
Obvyklé zastoupení |
|
Celulóza |
50% |
|
Hemicelulózy |
20% |
|
Lignin |
15% |
|
Bílkoviny |
6% |
|
Nižší sacharidy |
6% |
|
Pektin |
1,5% |
|
Vosky a pigmenty |
1,5% |
Složení se pochopitelně mění podle toho, z které rostliny opad pochází. Např. v jehličí bývá více vosku. Pouze cca 12% opadu patří mezi snadno odbouratelné látky (bílkoviny a nižší sacharidy), ostatní složky jsou obtížně odbouratelné.
Rychlost odbourávání opadu záleží na mnoha faktorech, mezi nejdůležitější patří:
V našich podmínkách trvá rozklad listí obvykle 1-3 roky, jehličí 8-10 let (jehličí má na povrchu obtížně překonatelnou voskovou kutikulu). Zbytky dřeva (pařezy, padlé kmeny apod.) se mohou rozkládat i několik desítek let. V tropických lesích je rychlost podstatně vyšší, listy jsou odbourány za několik měsíců a prakticky veškerý opad (s výjimkou velkých kusů dřeva) je rozložen ještě v tom samém vegetačním období.
Strategie postupného odbourávání souvisí s dostupností živin:
Je třeba dodat, že v posledních fázích, kdy rozkladu dominují saprotrofní houby, se uplatňují i bakterie, které zneužívají extracelulárních houbových enzymů a odčerpávají houbě rozložené oligosacharidy a monosacharidy. Mnoho hub proto produkuje různé látky inhibující bakterie (např. antibiotika), kterými se snaží toto vykrádání zastavit.
Termínem humus je označována nejúrodnější část půdy. Jedná se o soubor převážně organických látek v různé fázi rozkladu. Humus je tvořen zejména humusovými látkami. Humusové látky mají kladný vliv na kvalitu půdy, zlepšují její strukturu, zvyšují absorpční schopnost pro vodu a minerální živiny, mají pufrovací schopnosti a udržují tak konstatnější pH a v neposlední řadě svou tmavší barvou zvyšují absorpci světla, což vede mj. ke zvýšení teploty půdy.
Chemickou strukturu humusových látek nelze přesně definovat. Jedná se o polymerní látky s různou molekulovou hmotností, převládajícími prvky jsou uhlík, kyslík a vodík, ale vyskytují se i dusík a síra. Nacházené strukturní motivy často připomínají lignin, obsahují hodně aromatických kruhů a alifatických řetězců, dále četné hydroxylové, alkoxylové a karboxylové skupiny. Disociované karboxyly tvoří často komplexy s ionty kovů.
Humusové látky dělíme podle rostoucí molekulové hmotnosti na několik typů:
Vznik humusových látek není dosud plně objasněn a existuje mnoho teorií, které ho vysvětlují. Pravděpodobně se ale na jejich tvorbě podílí mnoho různých procesů, ve kterých hrají důležitou roli i mikroorganismy a jejich extracelulární enzymy. Část humusových látek vzniká z nerozložených zbytků dřeva a opadu, zejména z neodbouraného, částečně odbouraného nebo chemicky modifikovaného ligninu. Tomu napovídá i podobná struktura huminových látek a ligninu. Část humusových látek vzniká pravděpodobně polymerací nízkomolekulárních látek, zejména působením extracelulárních oxidativních enzymů produkovaných mikroorganismy. Podíl na tvorbě humusových látek mají nepochybně i neenzymatické chemické reakce probíhající i bez účasti mikroorganismů.
Odbourávání humusových látek je velmi pomalé a je tím pomalješí, čím vyšší je molekulová hmotnost. Humusové látky obvykle neslouží jako zdroj uhlíku a energie, byť by představovaly velice bohatý zdroj. To evokuje hypotézu, že rozklad humusových látek probíhá spíše neúmyslně, jen jako vedlejší produkt působení extracelulárních enzymů. Vzhledem k nedefinované proměnlivé struktuře se uplatňují zejména málo specifické enzymy, např. lignolytické (lakáza, peroxidázy, oxidázy). Působení těchto enzymů ale může vést i k opačnému procesu, tedy k polymeraci nízkomolekulárních látek. Při rozkladu jsou molekuly humusových látek štěpeny náhodně na různých místech a postupně se snižuje jejich molekulová hmotnost. Při analýze pak zjišťujeme postupné snížení podílu huminů a huminových kyselin a naopak zvýšení podílu fulvokyselin.
Soužití nejrůznějších mikroorganismů s rostlinami je velice časté. Nejvíce interakcí probíhá v oblasti kořenů, kde vymezujeme tzv. rhizosféru. Ta představuje oblast půdy, která je bezprostředně ovlivněna kořeny.
Rhizosféra představuje druhoě bohaté společenstvo mikroorganismů (zejména bakterií, hub a prvoků) a makroorganismů (kromě kořenů rostlin tu nacházíme běžnou půdní faunu, např. hlístice či hmyz). V rámci společenstva existuje mnoho vzájemných vztahů. V rhizosféře bývá obvykle zvýšená koncentrace organismů a zejména mikroorganismů. Důvodem je zejména větší nabídka živin využitelných mikroorganismy. Jedná se např. o odumřelé části kořenů, ale i o látky produkované kořeny do prostředí, zejména nízkomolekulární zdroje uhlíku. V rhizosféře vznikají četné potravní řetězce, ve kterých mikrooorganismy hrají jak roli konzumenta (např. rozkládají nejrůznější odpadní látky), tak roli potravy (pro prvoky či vyšší organismy).
Rhizosféra působí na rostlinu obvykle celkově pozitivně, zejména proto, že přítomné (mikro)organismy obohacují půdu např. zejména o vázané minerální živiny. Pozitivně také působí bohatost společenstva. Do bohatého společenstva mají další mikroorganismy ztížený vstup, což představuje pro rostlinu např. ochranu proti vnějším patogenním MO. Mnohé procesy probíhající v rhizosféře (např. odbourávání odumřelých buněk) nemají pro rostlinu žádný význam. Rhizosféra může působit na rostlinu i negativně. I když je průnik patogenních mikroorganismů ztížen, přesto stále mohou do konsorcia proniknout, usadit se v něm a rostlině škodit.
Některé rostliny zejména luskoviny (hrách, fazole, jetel, sója apod.) žijí v zajímavé symbióze s bakteriemi rodu Rhizobium[1]. Bakterie žijí přímo v kořenech rostlin, kde je rostlina zásobuje výživou (organickými kyselinami a přiměřeným množstvím kyslíku). Bakterie naoplátku fixují vzdušný dusík a produkují amonné ionty přímo do kořene. Fixace dusíku je energeticky náročný proces a volné bakterie ho obvykle neprovádí. Redukci dusíku na amonné ionty zajišťuje enzymový nitrogenázový komplex, který je citlivý na kyslík. Pravděpodobně i proto izoluje rostlina bakterii v kořenech a dávkuje jí kyslík.
Limitaci kyslíkem využívá rostlina i jako mechanismus pro trestání „neposlušných“ bakterií. Může se stát, že bakterie zneužije rostlinných živin, přestane fixovat vzdušný dusík a ušetřenou energii věnuje nekontrolovanému buněčnému dělení. Rostlina měří množství produkovaného amoniaku a málo výkonné bakterie omezuje kyslíkem. „Neposlušné“ bakterie tím sice přímo nezlikviduje, ale značně omezí jejich potomstvo. Rostlina tak vlastně ovlivňuje evoluci směrem k větší symbióze a závislosti bakterie na rostlině.
Mykorhíza[2] představuje symbiózu vláknitých hub s kořeny rostlin. Nemusí jít vždy o mikroskopické houby, mykorhíza je běžná u vyšších hub, které se běžně sbírají k jídlu. Odhaduje se, že mykorzhízy je schopno až 95% rostlinných druhů a je tedy velice rozšířená.
Mykorhíza představuje oboustranně prospěšné soužití. Houbové mycelium funguje fakticky jako prodloužení rostlinných kořenů, přičemž toto prodloužení je mnohonásobné ve srovnání s kořenem rostliny. Zejména mycelium lesních hub může mít délku i v kilometrech a může fungovat fakticky jako „potrubí“ a transportovat živiny z jednoho konce lesa na druhý. Houbové mycelium umí lépe přijímat z prostředí minerální látky a vodu a ty dodává rostlinně. Kromě toho se projevuje i ochranná funkce mycelia – kořen obalený houbovým myceliem je hůře napadnutelný parazity či patogenními mikroorganismy. Přítomnost houby navíc stimuluje život v rhizosféře, z čehož rostlina obvykle těží. Houba jako heterotrofní organismus naoplátku získává od rostliny uhlíkaté látky (zejména sacharidy), kterých má rostlina jako autotrof obvykle přebytek.
Je mnoha pokusy prokázáno, že po uskutečnění mykorhízy se oběma symbiontům lépe daří. Např. ve školkách vysazované stromky mají v přítomnosti houby větší pravděpodobnost uchycení a také rychleji rostou. Podobně i houba rychleji roste při soužití s rostlinou. Mnohé houby jsou pak na rostlině úplně závisle a nejsou schopné dlouhého přežití bez kontaktu s kořeny. Typické je to právě pro velkou část sbíraných hub, např. rod Leccinum (zahrnující mj. oblíbené houby křemenáče a kozáky) je schopen růstu jen v kontaktu s kořeny některých rostlin, např. břízy. I proto jsou tyto houby nacházeny téměř výhradně pod břízami. Úplná závislost rostliny na houbě nebývá obvyklá.
Z hledisky způsobu kontaktu houbového mycelia s kořenem rozeznáváme tzv. endomykorhízu a ektomykorhízu. Při endomykorhíze proniká houbové mycelium až mezi buněčnou stěnu a cytoplazmatickou membránu rostlinných buněk. Tento typ, označovaný často také jako arbuskulární mykorhíza, je nejčastější (cca 85% rostlin). Ektomykorhýza představuje stav, kdy houbové mycelium neproniká pod buněčnou stěnu kořenových buněk, ale ve většině případů pouze obaluje kořen. Tento typ mykorhízy je znám u přibližně 10% rostlin, převážně u dřevin, a má proto značný hospodářský význam.
Biofilm je společenstvo mikroorganismů tvořících odolnou slizkou vrstvu na pevném podkladu. V biofilmu je vysoká hustota mikrobiálních buněk spojených vhodným pojivem. Pojivo je tvořeno nejčastěji polysacharidy (např. dextrany) a bílkovinami. Biofilm může být tvořen jedním druhem mikroorganismu nebo se může jednat o společenstvo mikroorganismů V jednom biofilmu mohou být pohromadě i zástupci různých domén, např. bakterie, prvoci a archea.
Význam biofilmu je zejména ten, že představuje pro zúčastněné mikroorganismy dodatečnou ochranu. Mikroorganismy sdružené do biofilmu jsou obecně odolnější než volné mikroorganismy. To má mnoho negativních dopadů zejména na člověka a jeho činnosti. Některé patogenní mikroorganismy tvoří biofilmy v těle. Zatímco volné mikroorganismy jsou prakticky neškodné, tvorbou biofilmu se aktivuje jejich patogenní působení. Odolný biofilm je navíc těžké zničit, chrání mikroorganismy jak před imunitním systémem, tak před účinkem léčiv. Antibiotika často nezabírají ani ve vysokých dávkách. Zubní plak také není nic jiného než biofilm mikroorganismů. Při nedostatečném čištění se může tvořit biofilm na kontaktních čočkách a následně způsobovat oční infekce. Biofilmy mikroorganismů se také mohou tvořit v různých zařízeních, např. v potrubí, klimatizacích, na vlhkých povrchách apod. Tvorby biofilmu naopak využívají tzv. biofilmové reaktory (např. při čištění odpadních vod). V takovém případě se vhodný povrch nechá záměrně zarůst biofilmem a takto imobilizované mikroorganismy pak provádějí příslušné žádoucí přeměny (např. rozklad organických látek z v odpadní vodě).
Vznikem biofilmu se mikroorganismy dostávají do poněkud odlišného fyziologického stavu. Obvykle se v biofilmech uplatňuje quorum sensing, tedy fyziologická odpověď mikroorganismů na zvýšenou koncentraci buněk. Buňky začnou produkovat pojivo a své chování a metabolismus podřizují celému společenstvu. Např. buňky v kontaktu s podkladem se snaží na tento co nejlépe upevnit, naopak buňky na povrchu plní spíše obranné funkce.
V biofilmech je vysoká koncentrace mikrobiálních buněk a díky tomu dochází k diferenciaci metabolismů. Na buňky v hlubších částech vrstvy se obvykle nedostanou všechny živiny z povrchu, protože jsou zkonzumovány výše umístěnými organismy. Běžné tak je, že povrchové mikroorganismy provádějí aerobní respiraci, zatímco mikroorganismy ve spod už žádný kyslík nemají a nezbývá jim než fermentovat nebo provádět anaerobní respirace. Spodní buňky také často využívají odpadních produktů vyšších buněk a obvykle mají nižší metabolickou aktivitu.
Mikroorganismy se vyskytují prakticky všude a tedy i ve vzduchu. Množství mikroorganismů ve vzduchu ale není velké. Mikroorganismy ve vzduchu nejsou obvykle volné, ale bývají vázané na pevné částice, zejména prachové, popř. jsou součástí kapének tekutin. S tím souvisí i jejich výskyt v různých prostředích, více organismů je tam, kde se více práší. Ve volné přírodě proto bývá obvykle méně vzdušných mikroorganismů ve srovnání např. s rušnou ulicí, či průmyslovou oblastí. Podobně nad mořem je méně mikroorganismů než nad pevninou. V uzavřených místnostech se nachází méně mikroorganismů než venku.
Zamyslete se nad lidským tělem jako ekosystémem obývaným mikroorganismy a definujte hlavní faktory ovlivňující výskyt a aktivitu mikroorganismů (kožní mikroflóra, střevní mikroflóra, vaginální mikroflóra, ústní mikroflóra atd.). Nápověda - významnými faktory jsou např. živiny, dostupnost vody, teplota, odstraňování, fyziologické cykly apod.
Zamyslete se nad rozdílem v půdní mikroflóře u zemědělských orných a neoraných půd.
Popište základní ekologické vztahy mezi organismy a ukažte je na příkladech.
[1] Podle nejnovějších výzkumů, založených zejména na analýze genetických sekvencí, byl rod Rhizobium rozdělen na několik dalších rodů. Jako Rhizobia jsou nově nazývány všechny bakterie schopné fixovat vzdušný dusík v symbióze s rostlinou.
[2] Z latinského myces = houba a rhiza = kořen
Podle odhadů biologů poznal člověk většinu nebo alespoň více než 50% živočišných i rostlinných druhů. U hub se ale předpokládá poznání jen cca 20%, u bakterií a archeí je pak toto číslo řádově menší (řádově jednotky %).
Nejkompletnější přehled bakterií a archeí a včetně jejich taxonomického zařazení je …. let je publikováno v Bergey’s maual of systematic bacteriology. Toto kompendium vychází v pravidelně aktualizovaných vydání a zohledňuje nejnovější výzkumu. Kromě manuálu systematické bakteriologie je publikován i zjednodušený Bergey’s manual of determinative bacteriology zaměřený na praktické určování bakteriálních rodů a druhů.
Zatímco nejnovější vydání Bergeyova systematického manuál zavádí zcela nový taxonomický systém bakterií založený na jejich fylogenezi, zjednodušený určovací manuál se stále drží praktičtější systému, který je založen zejména na fenotypových vlastnostech bakterií. Nový systém dělí doménu Bacteria na 23 kmenů. Dosud používaný systém pak uvádí 37 skupin bakterií podle jejich fenotypové podobnosti. Oba taxonomické systémy se prolínají, mnohé nově definované kmeny zahrnují mikroorganismy z různých skupin starého systému, naopak organismy řazené do jedné skupiny starého systému jsou někdy nově zařazeny do více kmenů. Vzhledem k praktičnosti staršího systému zejména pro určování bakterií, je tento systém stále používán a bude použit i v následujícím textu.
Za zmínku stojí následující fakta vyplývající
z bakteriální evoluce:
Vysvětlete základní taxonomické skupiny mikroorganismů v kontextu evoluce.
Vysvětlete rozdíl mezi fenotypovým a fylogenetickým taxonomickým systémem, pokuste se definovat hlavní výhody a nevýhody.
Obor
limnologie (hydrobiologie) jako vodní ekologie, paleolimnologie;
fyzikální a chemické
vlastnosti vody, pohyby vod
- charakterizujte limnologii jako ekologickou disciplínu a vymezte její hlavní součásti
- jaký je význam Ženevského jezera ve světové limnologii?
- uveďte základní vlastnosti vody a jejich význam pro život ve vodách
- vysvětlete princip a mechanismus míchání nádrží a jeho význam
- zmiňte kategorie jezer podle typu míchání
- vysvětlete vznik Langmuirovy cirkulace
- co jsou to séše (seiches)?
- popište intenzitu pronikání světla pod vodní hladinu
- jaký je význam světelných poměrů pro vodní ekosystémy?
- charakterizujte eufotickou vrstvu nádrže
- jaký je princip měření průhlednosti Secchiho deskou a jaké má toto měření využití v rybářské a rybníkářské praxi?
- jak vzniká teplotní stratifikace nádrží a jaký je její význam pro chemismus a oživení?
- popište možné typy stratifikace kyslíku na profilu nádrže
Dusík se v přírodě vyskytuje v mnoha formách. Nesloučený dusík (N2) tvoří cca 78% vzduchu a je tedy velmi rozšířený. Ve vzduchu dále najdeme v malém množství anorganické oxidy dusíku (zejména NO a NO2), které se do něj dostaly činností člověka. Anorganické formy dusíku v přírodě zahrnují zejména dusičnany, dusitany a amonné sole. V organických látkách zejména biologického původu má dusík nezastupitelné místo. Je součást bílkovin, nukleových kyselin, biogenních aminů a mnoha dalších látek nacházených v živých soustavách.
Na přeměnách nejrůznějších forem dusíku se podílejí rostliny, živočichové i mikroorganismy, přičemž většinu přeměn ale umí právě jen mikroorganismy (viz obrázek). Živočichové přijímají jen organický dusík, jako odpadní produkt produkují nejčastěji amoniak (často ovšem převedený na méně toxické odpadní formy, např. močovinu) popř. některé formy organické dusíku, např. kyselinu močovou. Rostliny naopak přijímají jen anorganický dusík, buď v podobě amonných iontů nebo dusičnanů. Výjimkou jsou parazitující rostliny, které ze svého hostitele přijímají i hotové dusíkaté organické látky a také masožravé rostliny, které využívají organický dusík svých obětí.
Ostatní přeměny jsou doménou mikroorganismů (viz obrázek).
Amonifikace je proces, při kterém jsou uvolňovány amonné ionty z organických látek. Tyto procesy nejsou u mikroorganismů příliš obvyklé, protože mikroorganismy disponují velice ekonomickým metabolismem. Zbavování se organického dusíku, který by mohli úsporněji využít ve vázané podobě, tak představuje v podstatě plýtvání. S amonifikací se proto setkáváme jen tehdy, roste-li kultura na substrátu, ve kterém je přebytek dusíku nad uhlíkem. Druhý případ je tehdy, není-li organický substrát přímo zabudovatelný do buněk mikroorganismů a proto je ho třeba dočasně rozložit. Uvolněné amonné ionty jsou pak ale znovu asimilovány.
Asimilace amonných iontů je opakem amonifikace. Tento proces provádí naprostá většina mikroorganismů, mají-li amonné ionty jako zdroj dusíku. Jsou-li v prostředí dostupné hotové aminokyseliny, dává většina mikroorganismů přednost jim před amonnými ionty, protože na jejich zabudování do svých buněk spotřebují méně energie. Amonné ionty jsou naopak preferovány před ostatními využitelnými formami dusíku.
Asimilace dusičnanů a dusitanů je proces, při kterém mikroorganismy (zejména bakterie) zabudovávají dusičnany a dusitany ve formě aminoskupin do svých organických látek. Většina bakterií je schopná tento proces provádět. Asimilace probíhá několikastupňovou redukcí (dusičnanyà dusitanyàamonné iontyàorganické látky) a vyžaduje dodání energie a redukčních činidel. Proto většina bakterií dává přednost využívání organických látek a fixaci amonných iontů před asimilací dusičnanů a dusitanů.
Nitrifikace je proces oxidace amonných iontů na dusitany a v dalším kroku až na dusičnany pomocí kyslíku. Provádí ho tzv. nitrifikační bakterie, které tímto způsobem získávají energii, jedná se tedy o lithotrofní organismy. Tento proces má velký význam při čištění odpadních vod, protože amoniak (resp. amonné ionty) jsou v odpadních vodách hojně zastoupené. Nitrifikaci provádějí např. rody Nitrobacter, Nitrosomonas či Nitromonas.
Denitrifikace je proces rozkladu dusičnanů a dusitanů na redukovanější formy dusíku, zejména na plynný dusík N2. Jedná se proces, který nevyžaduje kyslík a kyslíkem je obvykle reprimován. Jsou nicméně známé i bakteriální rody, které denitrifikaci provádí i v přítomnosti kyslíku. Při těchto procesech slouží dusičnany a dusitany jako oxidační činidla pro organické látky a organismus tak získává energii. Proces probíhá podobně jako aerobní respirace, jen konečným akceptorem elektronů v dýchacím řetězci není kyslík, ale právě dusičnany či dusitany. Protože kyslík je silnější oxidační činidlo, dává většina mikroorganismů přednost aerobní respiraci před touto anaerobní (tzv. kyslíkový efekt viz kapitola…). Denitrifikace má význam při čištění odpadních vod jako doplněk nitrifikace. Kombinací obou procesů je poměrně jedovatý amoniak převeden až na neškodný plynný dusík. Denitrifikaci provádí zejména heterotrofní bakterie, často gramnegativní, např. rody Proteobacteria, Pseudomonas, Alcaligenes, Thiobacillus atd.
Fixace vzdušného dusíku je proces, při kterém je dusík ve formě N2 redukován až na amoniak a obvykle zabudován do organických látek buňky. Protože plynný dusík N2 je termodynamicky velice stabilní molekula, vyžaduje tento proces dodání velkého množství energie (12-16 ATP na fixaci jedné molekuly N2). Tento proces má klíčový význam v uzavření koloběhu dusíku, protože jinak by se většina dusíku postupně přeměnila na plynný dusík. Tento proces provádí jen některé skupiny bakterií. Rod Rhozobium fixuje dusík v symbióze s luskovinami (viz kapitola 9.5.1) a má tak velký význam pro úrodnost půdy. V symbióze s některými rostlinami dále fixují vzdušný dusík i některé aktinomycety (vláknité půdní bakterie), např,. rod Frankia. Nesymbioticky (pro sebe) fixuje vzdušný dusík rod Azotobacter a také Cyanobacterie (sinice).- co je to specifická vodivost vody?
- popište pufrační systém hydrogenuhličitan – kyselina uhličitá
- uveďte příčiny a důsledky eutrofizace, možnosti nápravy
- uveďte příčiny a důsledky antropogenní acidifikace, možná řešení
- jaká jsou hlavní společenstva stojatých vod? – uveďte zástupce
- jaká jsou hlavní společenstva tekoucích vod? – uveďte zástupce
- jak jsou vodní organismy přizpůsobeny životu v prudce tekoucích vodách?
- jaké jsou způsoby dýchání vodních organismů a co jsou „fyzikální plíce“?
- jaké jsou adaptace živočichů na nízké množství kyslíku ve vodě?
- uveďte příklady různých způsobů získávání potravy u vodních bezobratlých
- jaký je rozdíl mezi sinicemi a řasami?
- co je to tzv. summer kill a winter kill?
- představte základní ekologické typy vodních makrofyt, s příklady
- uveďte příklady nepohlavního rozmnožování a trvalých přežívacích stadií vodních bezobratlých
- jaké jsou základní ekologické skupiny vodních bezobratlých podle způsobu pohybu ve vodě?
- jaké jsou základní ekologické skupiny vodních bezobratlých podle způsobu získávání potravy?
- jaké typy organismů můžeme očekávat v oligotrofních a naopak silně eutrofních vodách?